|
|||||||
Cruzamiento de bovinos lecheros Alberto Valle*
|
 |
||||||
|
Venezuela es un país tropical, donde gran parte de su
territorio presenta altas temperaturas ambientales y humedades relativas; topografía
irregular, escasa fertilidad del suelo y una precipitación muy variable. Bajo estas
condiciones resulta desfavorable explotar razas lecheras importadas, debido a los
trastornos fisiológicos de adaptación al clima tropical y como consecuencia de ello, la
disminución en la producción de leche, retardo en el crecimiento, alteraciones en su
ciclo reproductivo y, en general, un escaso retorno del capital invertido. Las razas nativas o criollas, aunque bien
adaptadas a estas condiciones tropicales, muestran poca precocidad y baja producción de
leche, lo que las hace ser poco productiva. Pero constituyen un valioso material genético
para la formación de rebańos lecheros para el trópico americano. Desde el punto de vista zootécnico, el
ganado criollo de Venezuela tiene su origen en diferentes razas ibéricas, entre las
cuales podemos mencionar las siguientes: gallega, barrenda, pirenaica, tudanca, andaluza y
castellana, con especial énfasis en estas dos últimas, donde al trascurrir de los ańos
fueron adquiriendo las características impuestas por el medio ambiente, donde por la
actuación de la selección natural muchas generaciones la transformaron en un grupo
étnico bien adaptado y que se reproduce con cierta facilidad. Sin embargo, tanto las razas especializadas
como las autóctonas representan separadamente una explotación poco rentable, por lo que
se deben incrementar los niveles productos de los bovinos bien adaptados a las condiciones
tropicales, pero respetando en lo posible los principios fundamentales del mejoramiento
genético. Principios del mejoramiento genético El aumento en escala regional y/o nacional
de la capacidad productiva de todo tipo de bovino deberá basarse, en la mayoría de los
casos, en reproductores procreados por un escaso porcentaje del número total de bovinos
del mismo tipo, dentro de un número relativamente pequeńo de explotaciones. Aceptado lo anterior, se debe reconocer que
todo mejoramiento de la capacidad productiva del ganado, dependerá con el transcurrir del
tiempo, de los continuos mejoramientos genéticos de los reproductores. Además, la cría de los reproductores
deberá estar dirigida al mejoramiento efectivo de la capacidad productiva, si se desea
que los resultados de esa procreación beneficie a la industria lechera como actividad
comercial. Tendiendo en consideración estos
principios, el aumento de la prdoducción de leche en el trópico solamente puede
efectuarse a través de tres métodos generales. Métodos generales 1. Mejoramiento dentro de los tipos de
bovinos ya existentes. Este método presenta la ventaja de aprovechar toda la
adaptabilidad a las condiciones locales y evita los costos de selección e importación de
bovinos de otras latitudes, con el riesgo de introducir enfermedades y problemas de
adaptación. Su inconveniente lo representa un progreso genético muy lento. 2. Obtención de nuevos tipos utilizando
animales de una raza, la cual ha sido mejorada únicamente hasta el grado necesario para
lograr un animal más productivo. La ventaja es un rápido mejoramiento inicial, obtenido
mediante inseminación artificial de un considerable número de vacas con semen de un
reproductor valioso. La desventaja de este método consiste en problemas de adaptación y
sus consecuencias. 3. Creación de un nuevo tipo de ganado,
mediante el cruzamiento sistemático de dos tipos distintos para combinar características
deseables. La ventaja está en asociar producción y adaptabilidad y su desventaja radica
en que debe ser efectuado en gran escala si se desea obtener resultados satisfactorios en
un tiempo razonable. Tanto el segundo como el tercer método
implican el cruzamiento de bovinos, diferenciándose solamente en que en el segundo se
obtienen animales de mercado, los cuales deberán ser renovados constantemente, mientras
que en el último método se están sentando las bases para la formación de una nueva
raza. En ambos se está aprovechando el principio genético de la heterosis. Heterosis El vigor híbrido o heterosis es la
superioridad individual de animales o plantas, que se obtiene por el apareamiento o cruce
entre progenitores menos relacionados entre sí por encima de los representantes promedio
de la población de su procedencia. El apareamiento de tales progenitores se
denomina exogamia y sirve, tanto para aumentar el vigor del animal descendiente como para
uniformizar el comportamiento zootécnico de la generación. En todas las poblaciones
existen individuos con características superiores e inferiores, cuya presencia provoca la
heterogeneidad dentro de la misma. La hexogamia disminuye las diferencias individuales y
tiende a uniformizar el comportamiento de la población. El máximo efecto de la exogamia se observa
al inicio de su práctica, o sea, en las primeras generaciones cruzadas y disminuye en las
generaciones posteriores. Su utilidad práctica reside en la dominancia de los genes
portadores de los caracteres favorables sobre sus pares (alelos), que aportan las
expresiones menos deseables del mismo caracter. Por lo tanto, la exogamia mejora el
comportamiento de los descendientes, pero disminuye la potencialidad genética de los
individuos sobresalientes en los cuales inicialmente los genes favorables se presentan en
forma de parejas uniformes (homocigotos). La aparición de un gene desfavorable,
heredado del otro progenitor, transforma el par de alelos en heterocigotos, repercutiendo
en el fenómeno de que el futuro reproductor va a proporcionar a la mitad de sus
descendientes las características menos deseables. La máxima práctica de la exogamia reside
en la producción de vegetales y animales para el mercado, bien sea para consumo o para
exposiciones, mientras que su repercusión sobre el mejoramiento genético a largo plazo
será reducida y a veces hasta negativa. Cruzamiento. Experiencia con bovinos Las pocas experiencias de exogamia con
bovinos han sido encaminadas principalmente para la determinación de las diferencias
entre razas y muy poco para conocer si el cruzamiento repercute en el éxito comercial. El
cruzamiento, como cualquier otra forma de exogamia, tiende a disminuir la potencialidad
genética del animal, haciéndolo más heterocigotos, reduce la eficiencia de la
selección entre los descendientes F1 (50% R1 - 50% R2) y mejora el mérito fisiológico o
productivo individual, debido a la dominancia general de los genes favorables para el
tamańo, vigor, fertilidad, adaptación, etc. Cuando los productos del cruzamiento son
utilizados para fines reproductivos, sus descendientes serán genéticamente más
variables que ellos y generalmente tendrán un promedio de potencialidad hereditaria de
valor más bajo. Si ambos padres son cruzados con 50% de genes R1 y 50% de R2, su prole
tendrá usualmente un valor genético medio individual inferior al de los abuelos. Muchas
veces la distribución de los descendientes de padres 50% de R1 - 50% R2 es asimétrica,
existiendo pocos individuos por encima del promedio de los padres, ubicándose la mayoría
por debajo de este. Además de la dominancia general de los
genes de efectos favorables, es probable que mucha de esta asimetría en la distribución
de los descendientes sea causada por interacción génica. Sin embargo, se ha observado
que algunas líneas híbridas presentan una buena capacidad de combinación (habilidad
combinatoria general) con la otra población en la cual va a ser utilizada. La heterosis no ocurre uniformemente en
todos los cruzamientos. Ni todas las razas, ni todos los animales dentro de la misma raza
se "combinan" igualmente bien, lo que se debe a la habilidad combinatoria
específica. La heterosis obtenida por el cruzamiento de
razas de animales domésticos, en la mayoría de los casos no sobrepasa de 2 a 8% de
aumento sobre el promedio de las razas usadas como progenitores en características como
el tamańo, la ganancia de peso, la fertilidad y otras de compleja fisiología. Ella es
generalmente máxima para vitalidad, medida por el porcentaje de hijos sobrevivientes
dentro de los procreados. No existen observaciones en mejoramiento
animal que se podrían comparar con el grado de heterosis que los geneticistas de maíz
obtienen frecuentemente al cruzar dos linajes endógamos. Aunque los principios genéticos
básicos son los mismos, aparentemente no existe en las razas de animales domésticos la
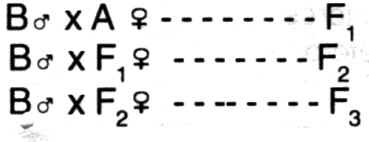
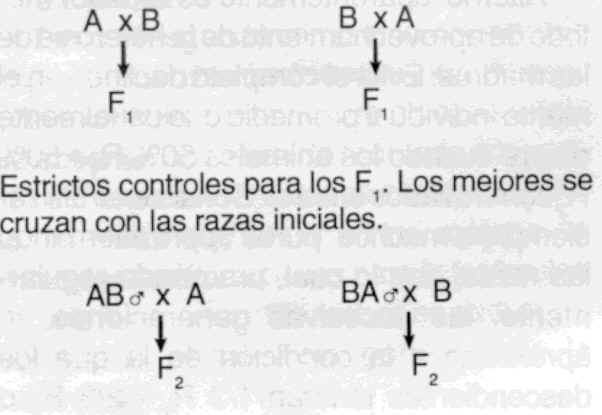
habilidad combinatoria comparable a los linajes puros de maíz. Tipos de cruzamiento - Industrial o de primera generación:
generalmente utilizado para obtener animales para la venta. A x B --------- F1 (mercado) Ventajas: - Absorbente (grading up o top
crossing):
utilizado para sustituir una raza por otra. - Alterno: aparentemente es el mejor
método de aprovechamiento de la heterosis de las madres. Evita el completo declineo en el
mérito individual promedio que usualmente ocurre cuando los animales 50% R1 - 50% R2 son
cruzados entre sí. Consiste en utilizar siempre a machos "puros", pero
alternando las razas, con lo cual, practicado regularmente, las sucesivas generaciones se
aproximan a la condición en la que los descendientes tendrán 1/3 R1 - 2/3 R2 o
viceversa. - Alterno rotacional: se utilizan
machos de dos razas (A y B).
Nota: después de la tercera
generación (F3), el procentaje de A y B tiende a asumir los valores de 1/3 y 2/3 en las
generaciones impares y 2/3 y 1/3 en las pares. Ventajas: Aprovechamiento de la heterosis (si los
machos no son los mismos). Requiere mucha dedicación y controles. Ventajas: Ventajas: - De retorno (back crossing): es lo inverso
del cruzamiento absorbente. - De bimestizos: consiste en el cruzamiento
de los F1 y/o F2 para tratar de lograr una nueva raza. Ejemplo de ello son las razas
Carora (Venezuela), Pitangueiras (Brasil), Santa Gertrudis (USA), Siboney (Cuba) y Jamaica
Hope (Jamaica), etc.
Dominancia: la disminución del vigor debido al endocruzamiento, sugiere en forma general, que en los animales domésticos existen muchos genes recesivos que presentan efectos deletéreos sobre el vigor del animal, variando de levemente perjudicial hasta letal. Debido a que existen muchos pares de genes influenciando la expresión de los caracteres cuantitativos, algunas razas y linajes endocruzadas podrían ser recesivas para algunos pares de genes; es decir, cuando muchos pares de genes controlan un caracter, una raza puede ser homocigota dominante para algunos pares y homocigota recesiva para otros (AABBCCdd), mientras que en otra raza podría ser, respectivamente, homocigota recesiva y homocigota dominante para esos mismos pares (aaBBCCDD). Cuando son cruzados animales de las dos raza, la F1 (AaBBCCDd) será superior a ambos ancestrales para ese caracter en particular, teniendo al final un gene dominante en cada par. Siendo la dominancia responsable de la heterosis, podría ser teóricamente posible tomar la superioridad de un linaje simple para obtener individuos homocigotos dominantes para todos los pares de genes, y si cruzamos entre sí a los animales con este genotipo sus descendientes tendrán todos el mismo genotipo. Sin embargo, estos homocigotos dominantes serían difíciles de reconocer, debido a que son semejantes al heterocigoto en relacióon al fenotipo. Además, es muy posible que existan más de dos pares de genes para regular el caracter, lo que complicaría más la tarea de establecer los homocigotos dominantes. Así que, aunque teóricamente es probable, en la práctica resulta poco menos que imposible. Sobredominancia: este tipo de acción génica contribuye para la manifestación de la heterosis y, en la práctica, muchos pares de genes con acción sobredominante pueden afectar la misma característica, aunque los efectos de los diferentes pares pueden ser de idénticas proporciones generalmente unos tienen mayores efectos sobre otros. Con este tipo de acción génica sería imposible fijar el heterocigoto para una simple característica, debido a que la acción es enteramente dependiente de la heterocigosidad. El único medio de aprovechar la ventaja del vigor sería la de formar primeramente un linaje endocruzado y usar los homocigotos entre sí. Los animales resultantes son sometidos a pruebas, comprobando cual de ellos se cxombina mejor y que produzca mayor grado de heterosis en sus descenientes. La mejor combinación de los ancestrales deberá ser cruzada nuevamente para producir heterocigosidad individual (esto es lo que exactamente se hace en la producción de maíz). Posteriores cruzamientos solamente disminuirán la proporción de heterocigotos de la población, con el consiguiente declineo de la la heterosis, regresando la media poblacional al promedio de los ancestrales iniciales. Epistasia: existen muchas formas de acción génica apistática, aunque sus efectos sobre las características cuantitativas sean difíciles de ser medidos, debido básicamente a su complejidad. En la dominancia y sobredominancia el efecto heterótico es debido a la interacción de genes de la misma serie alélica, siendo que muchos pares de alelos pueden afectar una misma característica. En la epistasia, la interacción es entre pares de genes no alélicos y en ella teóricamente posible fijar los efectos de la heterosis en un linaje simple, aunque en la práctica es extremadamente difícil, o lo que es lo mismo, es imposible mantenerla por medio de cruzamientos de individuos con alto grado de heterocigosidad. Existen otros tipos de interacciones inter-alélicas con resultados heteróticos y hasta algunos, teóricamente, pueden ocasionar una heterosis negativa. Afortunadamente la mayoría de los genes con efectos no deseables son recesivos, existiendo sin embargo ejemplos dominantes en animales de laboratorio. En todo caso, la heterosis máxima dependería de la presencia de no menos de un dominante para un máximo de pares alélicos y, teóricamente, podría ser posible seleccionar razas o linajes puros que tuviesen todos los genes dominantes en homocigosis para obtener su máxima expresión. Si nos concediéramos la posibilidad teórica de de lo anterior, tenemos que reconocer también que el mejoramiento de cada raza o linaje sería lento, debido a que existen grandes números de pares de genes que afectan los caracteres productivos. Por ejemplo, si solamente siete pares de genes son heterocigotos, podemos esperar 2,187 combinaciones diferentes; si son diez, esperaremos 59,049. Es decir, si dos animales son heterocigotos para n pares de genes, teóricamente pueden producir 3n tipos de descendientes. |
|||||||