|
1 * FONAIAP Estación Experimental Yaracuy. Actualmente en ICI Venezolana S.A. ** Universidad Central de Venezuela Facultad de Agronomía. Instituto de RECIBIDO: marzo 13 1989 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
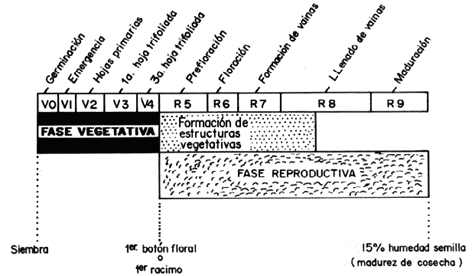
RESUMENP.C.: Andrector, Gynandrobrotica, fluctuación poblacional fenología. colonización emigración Phaseolus INTRODUCCIONLos coquitos perforadores (Coleoptera: Chrysomelidae), después del complejo Empoasca (Homoptera: Cicadellidae), son considerados las plagas de más amplia distribución y mayor importancia de la caraota y otras leguminosas comestibles en América latina (8, 12), lo cual es válido para nuestro país.1 Existen varios géneros de crisomélidos, específicamente de la subfamilia Galerucinae que atacan leguminosas comestibles en Venezuela, entre ellas están Andrector, Diabrotica y Gynandrobrotica. Las especies más importantes son A. arcuatus, A. ruficornis y G. equestris 2 (7), éstos son muy frecuentes en la región centro-norte del país (17) en la cual se llevó a cabo el presente estudio. Durante la fase larval se alimentan de raíces y nódulos nitrificantes, mientras que los adultos lo hacen del follaje y órganos de reproducción tiernos (5,17). Además, son eficientes transmisores de virus (2, 3). Con respecto a estudios sobre fluctuaciones poblacionales de crisomélidos plagas de leguminosas en zonas tropicales, se puede decir que se ha investigado muy poco. Por lo general, este tema constituye un aspecto secundario dentro de otro estudio, entre ellos tenemos a los realizados en Diabrotica balteata y Cerotoma (=Andrector)fascialis (l5) y en A. arcuatus y A. ruficornis sobre caraota (17) así como en las tres especies bajo estudio sobre soya (4). MATERIALES Y METODOSCampos de cultivo. Los muestreos se realizaron en la Estación Experimental "Samán Mocho (UCV, Facultad de Agronomía), cercana a la población de Tacarigua en el estado Carabobo, durante los meses de septiembre, octubre y noviembre de 1983, y en el Campo Experimental del Centro Nacional de Investigaciones Agropecuarias (FONAIAP) entre Maracay y El limón, en el estado Aragua, desde noviembre de 1984 hasta mediados de enero de 1985. E1 primer campo tenía forma rectangular con una superficie de 2 160 m2 (60 m x 36 m) subdividida en nueve parcelas de 240 m2 (20 m x 12 m) con 12 hileras de cultivo cada una. El otro campo era completamente cuadrado de 1 296 m2 (36 m x 36 m) y subdividido en nueve parcelas de 144 m2 (12 m x 12 m) de 15 hileras. Ambos campos fueron sembrados con caraota (Phaseolus vulgaris L.) cv. 'Tacarigua'. Se les proveía riego cada vez que fuera necesario y se mantuvieron libres de malezas, tanto adentro como en los alrededores. En el campo sembrado en 1984 fue necesaria una aplicación de fungicida para controlar la roya de la caraota (Uromices phaseoli) durante las últimas semanas del ciclo de cultivo. Muestreo de adultos En cada uno de los campos se realizaron muestreos semanalmente entre las 9:00 y 11:00 am desde la primera semana después de la siembra hasta cuando el cultivo alcanzó su senectud. Durante cada fecha de muestreo se tomaban tres muestras al azar por parcela. En total se obtuvieron 27 muestras por semana, lo que se consideró un muestreo. La metodología de toma de muestras varió dependientemente de la edad del cultivo. Durante las dos primeras semanas se tomaron las muestras mediante observación visual directa de las plantas, hasta cuando tuvieron más de una hoja trifoliada. Este método ha dado buenos resultados en otras leguminosas (9, 16). A partir de la tercera semana se utilizó el muestreo con malla por el método de golpe lateral, el cual consiste en colocar la abertura de la malla paralela en la hilera, muy cerca a la parte superior de las plantas, mientras que con la otra mano se golpea varias veces el follaje, de manera que los insectos caigan dentro de la malla. Inmediatamente después, la malla se coloca en el área próxima contigua a ser muestreada y se repite la operación hasta completar la longitud de la unidad de muestreo. Este método presenta ciertas ventajas para el muestreo de coquitos galerucinos de la caraota, sobre otros métodos convencionales como el malleo simple y el malleo doble (18). La unidad de muestreo consistió en 3 m de hilera, en el case del método de malleo utilizado equivale a 10 mallazos contiguos. Cada muestra se introducía en una balsa de papal con su respectivo número e inmediatamente se engrapaba, las mismas eran transportadas dentro de una cava de anime y posteriormente congeladas, con la finalidad de meter a los coquitos y presentarlos. Luego eran contados, separados por especie, sexo y tipo de adulto (Teneral/Maduro), como lo señala YEPEZ GIL (17) Crecimiento y desarrollo del cultivo E1 crecimiento y desarrollo del cultivo de caraota donde se realizaron los muestreos se caracterizó de dos formas. La primera como variación del área foliar a lo largo de su ciclo de vida, y la otra mediante una escala creada por CIAT (1) para identificar las diferentes etapas del ciclo de la planta, de acuerdo a su morfología y cambios fisiológicos; ésta refiere las observaciones a estados de desarrollo , lo que permite conocer la fenología del cultivo Para estructurar la escala se tomaron en cuenta las dos fases fundamentales, vegetativa y reproductiva, que ocurren sucesivamente en el ciclo de la planta; dentro de estas fases se identificaron diez etapas delimitadas por eventos fisiológicos importantes (Figura1). A fin de tener una visión más completa de la fenología del cultivo, se adiciona la Fase de Senectud y se delimita desde el momento cuando comienza una acelerada defoliación natural de las plantas. Este evento fisiológico ocurre solamente en algunas variedades, entre ellas la utilizada en esta investigación.
A lo largo de la presentación y discusión de ]os resultados se nombra entre paréntesis la codificación de las etapas de cultivo que se encuentra en el esquema del CIAT. Por último, el tiempo de duración del cultivo, que es la variable independiente de las gráficas presentadas, es un parámetro utilizado para expresar el crecimiento del cultivo pero presenta la desventaja que el ciclo biológico de la caraota varía con el genotipo de la planta y con los factores ambientales. Para cumplir estos objetivos fue necesario tomar una planta por subparcela durante cada uno de los muestreos. Se cortaban a nivel del cuello y se introducía en balsas plásticas para que no perdieran su turgidez y dificultaran sus mediciones y observaciones. Fueron llevadas al laboratorio y se anotaban características como tipo y número de hojas, presencia de órganos reproductivos y de fructificación y su estado, lo que permitía clasificarla dentro de una de las etapas. Posteriormente a estas mismas plantas se les midió el área foliar con un medidor de área licor(D modelo l1 3000. En algunos cases se utilizó un índice que permitía conocer el área de la planta mediante la aplicación de la ecuación de regresión ATP = (Y = 1,775 + 2,2072X) para la variedad en estudio, donde ATP = área total de la planta, Y = área de la hoja (trifoliolo) y X =ancho máximo del foliolo central en cuestión. 1 Métodos estadísticos.
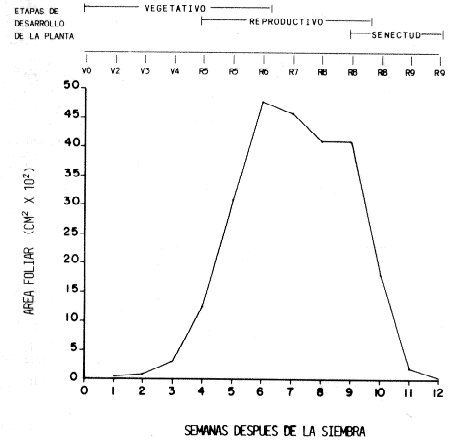
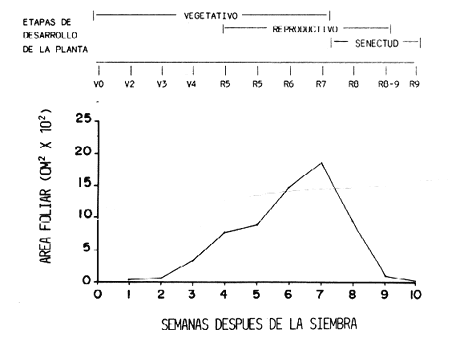
RESULTADOS Y DISCUSIONCrecimiento del cultivo. El cultivo sembrado en 1983 (Figura 2) presentó un crecimiento lento y constante durante las tres primeras semanas después de la siembra. Al formar las primeras hojas trifoliadas (V4), hubo una producción acelerada de follaje con un aumento sustancial del área foliar en un lapso de tres semanas (R6). Entre la cuarta y quinta semanas se solaparon los períodos vegetativo y reproductivo, ya que hubo producción de hojas y también de órganos de reproducción, debido a que el cultivar 'Tacarigua' es de crecimiento indeterminado. La fase reproductiva comenzó en la cuarta semana con la formación de botones florales (R5). En la sexta semana, cuando hubo máxima producción de follaje, ocurrió la floración (R6) que abarca casi tres semanas y se solapa en la última semana con la formación de vainas (R7) que se sucede principalmente en la séptima semana. Luego viene el periodo de llenado de vainas (R8), que es el más larga del ciclo y comienza en la octava semana. Poco después comienza el deterioro natural de las plantas, lo que se llamó Fase de Senectud, la cual se mantuvo a lo largo de las últimas tres semanas del cultivo. En este lapso ocurre la maduración de los granos (R9). El crecimiento de las plantas del campo de 1984 (Figura 3) se caracterizó por ser muy lento y con una reducción considerable del área foliar, aunque las etapas fisiológicas correspondientes se sucedieron cabalmente. En las tres primeras semanas hubo una producción de hojas de tamaño reducido y en poca cantidad; las plantas siguen esta tendencia de aumento lento en el área foliar por varias semanas, sin llegar a presentar ascensos marcados. La fase reproductiva comenzó durante la cuarta semana con la presencia de botones florales (R5) y en la sexta semana el cultivo se encontraba en plena floración (R6). Durante la séptima semana se obtuvo el máximo valor de área foliar, en este momento hubo un solapamiento de varias etapas ya que hubo mucha variación entre las plantas, tal vez por problemas de riego. En esta misma semana se detectó la presencia de roya, pero fue entre la octava y novena semanas cuando el daño en el follaje fue severa, ya cuando la planta se acercaba a su defoliación natural por senescencia y se encontraba en la fase de llenado (R8) y maduración de vainas (R9), estas etapas se acortaron un poco. Los muestreos se hicieron hasta la décima semana cuando ya quedaban muy pocos vestigios de follaje y habían madurado las vainas. Las diferencias de crecimiento expresadas como área foliar y semanas después de la siembra entre ambos campos, fueron bastante marcadas. El campo de 1983 produjo un follaje máximo promedio de 4 800 cm2, mientras que en el de 1984 solamente fue de 1 900 cm2, con una duración menor de dos semanas, lo que pudo ser debido a influencia de factores climatológicos y edafológicos diferentes en el campo sembrado en el CENIAP. Quizás la incidencia de roya aceleró el proceso. No obstante, hubo una tendencia semejante en ambos campos.
Los cambios fisiológicos se cumplieron de manera similar durante las dos ocasiones y se lograron delimitar bastante bien por la escala utilizada. Solamente durante las últimas etapas de la fase reproductiva se presentaron ciertas diferencias por las causas ya citadas. La caracterización del crecimiento del cultivo de esta forma es la que ha resultado de mayor importancia a la hora de estudiar las poblaciones insectiles, ya que permite realizar comparaciones de resultados procedentes de sitios diferentes y con distintos tiempos de duración, ya que se están caracterizando los estados de desarrollo. Comparación de valores promedio poblacionales entre especies. Las tres especies de coquitos perforadores se encontraron conviviendo en los dos lugares donde se llevaron a cabo los ensayos. Los promedios totales y el porcentaje que representa cada especie con respecto al total, para cada campo, se presenta en el Cuadro 1. Los valores promedios poblacionales de 1983 son significativamente diferentes entre especies. Lo mismo ocurre en el campo sembrado en 1984.
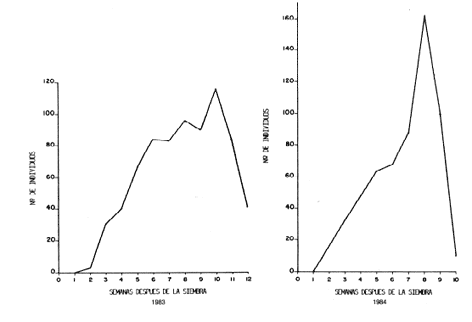
A. arcuatus es la especie con mayor densidad poblacional, seguida por A. ruficornis y, finalmente, G.. equestris. La dominancia de la primera especie con respecto a las otras dos pudiera ser debido a ciertos aspectos biológicos tales como menor duración de preadulto y mayor fecundidad, que le proporcionarían ciertas ventajas competitivas en el agroecosistema caraota (19). Los valores porcentuales de cada especie, obtenidos del total de promedios de la población, fueron convertidos en valores proporcionales. La proporción A. arcuatus A. ruficornis c. equestris quedó representada numéricamente con cifras aproximadas a 6: 3: 0,5, no obstante, que ambos campos tenían densidades poblacionales totales distintas y estaban completamente separadas en tiempo y espacio. Esta proporción es parecida a la obtenida por FELACO (4), en la Estación Experimental Samán Mocho, de 7:2:1 sobre soya para las tres especies, respectivamente. Fluctuaciones poblacionales. La información recopilada sobre fluctuación poblacional de los adultos de las tres especies estudiadas, es presentada como número total de individuos para cada muestreo . Para tener un conocimiento más de tallado de los cambios que ocurren en la población, se presenta además la información sobre los tipos de adulto (Teneral/Maduro) y el sexo. La discusión de la información para cada especie se presenta para los dos campos muestreados, simultáneamente. Es importante señalar que se hace especial énfasis y se discute en mayor detalle la especie A. arcuatus por ser la más abundante, luego A. ruficornis y, por último, G. equestris, en la que no se pueden discutir ciertos aspectos, dada la baja densidad observada. Durante la primera semana después de la siembra, cuando las plantas solamente presentaban el por de hojas cotiledonares, no se llegó a muestrear ningún individuo de los estudiados, posiblemente porque había pasado muy poco tiempo y el cultivo no estaba lo suficientemente atractivo para ser colonizado por un buen número de individuos o había una baja población vecina potencialmente invasora. Andrector arcuatus Olivier. La curve poblacional de .. arcuatus, para 1983 (Figura 4) se inició con un aumento sostenido de la población, debido a la entrada de nuevos individuos que colonizaron el cultivo' y que se acumularon posiblemente por la baja mortalidad en las primeras semanas de vida. Este proceso se observó hasta la sexta semana, a partir de la cual hubo una disminución de la tasa de incremento poblacional, lo que hizo, que la población se mantuviera a. altos niveles poblacionales con poco aumento a lo largo de cuatro semanas, y luego ocurrió el pica máximo de crecimiento, de la población, que como se explica más adelante está conformada en buena proporción por individuos que se han desarrollado dentro del campo y conforman la primera y única generación que se produce a lo largo del ciclo de cultivo de caraota.
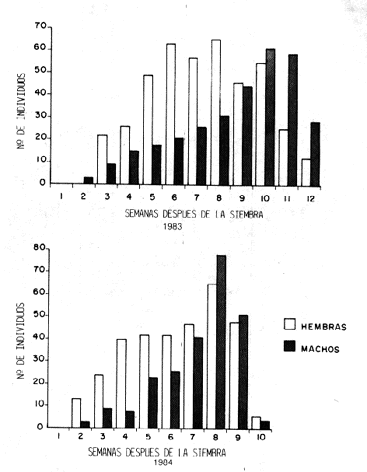
Esta sección de la curve, donde la población se mantuvo a niveles altos, conlleva a una situación que pudiera tener interés económico. En la primera de estas semanas, el cultivo ha acumulado su mayor cantidad de área foliar y durante las semanas siguientes se produjo gran cantidad de flores ((R6)) y se formaron las vainas (R7). Los coquitos, en busca de material tierno para su alimentación, consumen las hojas más jóvenes así como las flores y vainas en formación. Ya cuando ocurre el pica poblacional de adultos, el cultivo ha comenzado su deterioro natural (R8R9) y muchas vainas están llenas, sólo les falta secar. Posterior a este pica, se observó en la curve un descenso brusco del número de individuos de la población debido a que la producción de partes tiernas de la planta había cesado y la casi totalidad del follaje se encontraba seco. En este momento se produce la emigración de los individuos en busca de otros hospederos aptos para proveerles alimento y posibilidad de sobrevivencia a una nueva progenie. Para 1984 (Figura 4) se presentó una tendencia bastante similar a la del año anterior, aunque se observaron ciertas diferencias como ocurre normalmente en los procesos biológicos, además, hay variaciones geográficas, climatológicas, edafológicas, entre ambos campos, que van a influir sobre la entomofauna que se desarrolla a expenses de ellos. El período de crecimiento sostenido se presentó hasta alcanzar el pica máximo con un número de individuos mayor al de 1983. Después del pica poblacional viene un descenso drástico que en las próximas dos semanas lleva a la población a niveles muy bajos, además que el cultivo estaba completamente marchito por el severo ataque de roya que lo afectó en estas últimas semanas. La Figura 5 presenta a la población total dividida en sus componentes de tipo de adulto, o sea, en tenerales y maduros. Esta separación permite interpretar mejor los cambios que ocurren a lo largo del tiempo, así como detectar la aparición de la primera generación. Como puede observarse, en ambos cases se detecta la presencia de adultos tenerales en la sexta semana, es decir, cuatro semanas después que se muestrearon los primeros adultos colonizadores, lo que aproximadamente coincide con el tiempo de duración de la fase preadulta (huevo, larva, prepupa y pupa) obtenida en laboratorio (20). Para 1983 la curve de los tenerales sigue la misma tendencia ascendente que lleva la curve de maduros, pero no tan marcada, y en sólo cinco semanas llega a su pica máximo, lo que sería la décima semana de muestreo, coincidiendo con el pica máximo de adultos maduros. Un case muy similar se presenta al año siguiente donde la cuna de tenerales sigue un aumento sostenido muy parecido al de la cuna de adultos maduros y posteriormente coinciden ambos picas poblacionales. Es importante hacer notar que durante este año, la proporción de tenerales ocupa alrededor de un tercio de la curve total en la octava semana, lo que permite extrapolar que la mayor parte de la población pertenece a la descendencia de los colonizadores. Si se toma en cuenta que los adultos recién emergidos mantienen su carácter de tenerales alrededor de dos días, y que los muestreos estuvieron distanciados siete días, es difícil hallar una proporción alto de tenerales. Cuando se separó la población muestreada en sexos con la finalidad de ver como variaba la proporción hembras:machos a lo largo del ciclo de cultivo, fueron encontradas ciertas tendencias similares durante los dos años (Figura 6). Para 1983, desde el momento en que se detectan individuos en los muestreos, la proporción favorece significativamente a las hembras hasta la novena semana, cuando la proporción sexual se equilibra durante dos semanas y posteriormente se hace favorable a los machos hastíe el final del ciclo. Durante 1984 se presenta la misma situación, excepto que los machos no llegaron a predominar sobre las hembras después de la séptima semana cuando se establece la proporción 1:1 entre los sexos, situación que se mantiene hasta finalizar los muestreos. Aunque la proporción de machos no fue significativamente superior a las hembras, se observó la misma tendencia numérica en los dos muestreos anteriores al último (Cuadros 2 y 3). La proporción de sexos de los individuos tenerales no presentó diferencias significativas, tal como se observó en las crías de laboratorio (17), de lo que se desprende que el pica poblacional está formado por individuos emergentes más que por colonizadores. En resumen, se puede observar que durante 1983 se presentó una inversión de la dominancia de los sexos, que favoreció primeramente a la fracción femenina y después, en el momento de senescencia del cultivo, favoreció a los machos. Pero en 1984 no se cumplió el mismo proceso cabalmente, ya que los machos no llegaron a superar significativamente a las hembras. Andrector ruficornis Olivier. La curve poblacional de los adultos (Figura 7), muestreados durante el primer ensayo (1983), presentó un aumento sostenido hasta llegar al pica poblacional en la séptima semana, posteriormente descendió de igual manera. La gráfica obtenida para el ensayo sembrado en el Campo Experimental del CENIAP se inició muy parecida a la del año anterior, pero en las dos semanas siguientes al pica poblacional la población se mantuvo a altos niveles y luego vine el descenso característico hacia la etapa senil del cultivo. Con respecto a los individuos tenerales de A. rufircornis, durante 1983 se llegó a colectar un número muy bajo, específicamente en las semanas 10 y 11. Durante 1984 los tenerales se presentaron en la séptima semana de muestreo; en ambos años, la detección de tenerales se sucedió después del pico poblacional. La baja densidad de la especie no permite, con el sistema de muestreo utilizado, hacer mayores inferencias.
La proporción de sexos (Figura 8) durante el primer año de muestreo no mostró diferencias significativas durante las tres primeras semanas. Posteriormente, cuando la población aumentó, sí se reflejaron diferencias que favorecieron a las hembras hasta la séptima semana, cuando ocurrió el pica poblacional. En la semana siguiente se mantuvo la proporción 1:1 y en la novena semana pasó a favorecer a los machos. Durante las tres semanas siguientes la población no presentó ninguna diferencia significativa entre los sexos. Durante 1984 hay predominio bastante marcado de hembras durante las primeras cinco semanas, aunque la cuarta semana no mostró diferencias. En el resto de las semanas siguientes, a excepción de la penúltima, se observó equilibrio en la proporción hembras:machos. En general, durante las primeras semanas se observó una dominancia de hembras, posteriormente se equilibró y de ahí en adelante se mantuvo la proporción 1:1 (Cuadros 2 y 3). Gynandrobrotica equestris Fabricius. La densidad poblacional promedio de esta especie, como se dijo al principio, fue significativamente menor que las otras dos especies estudiadas. Como el número de individuos colectados fue muy pequeño se dificulta la consideración de los cambios que se presentaron en la curva. La información sobre esta especie se presenta en la Figura 9. La curve poblacional del primer campo mostró un paulatino aumento en el número de individuos y llegó a un máximo en la sexta semana; luego la población comienza a decrecer, no obstante, un ligero aumento observado en la octava semana. En el campo sembrado en 1984 la densidad fue aún más baja que en el año anterior, se detectaron los individuos en la cuarta semana y en la próxima siguiente se observó la máxima poblacional para luego declinar. En relación a la proporción sexual, durante 1983 hubo mayor proporción de hembras en la mayoría de los muestreos y en 1984 no hubo diferencia. En general, se observó una tendencia de aumento poblacional durante el desarrollo vegetativo del cultivo, hasta llegar a un pica máximo; en ambos años se asemejan a las curves de A. ruficornis. Posteriormente viene el descenso poblacional que es bastante gradual a diferencia de las dos especies de Andrector, Las curves poblacionales de las tres especies, en ambos campos, presentaron similitudes en sus tendencias, las cuales están íntimamente relacionada con la naturaleza temporal del hospedero. No obstante, solamente en las de A, arcuatus, por ser la especie más abundante, se observaron patrones bien definidos de dichos cambios. La interpretación de las variaciones poblacionales a lo largo de ambos ciclos permite dar una explicación general del proceso: la colonización del Fig. 8. Variaciones en el tiempo de sexos en adultos de A. ruficornis durante Fig. 9. Variación en el tiempo de la densidad y sexos de C. equestris durante 1983 y 1984. cultivo por parte de las diferentes especies se presentó muy temprano, cuando las plantas apenas mostraban sus primeras hojas y fue realizada principalmente por hembras. Durante las semanas siguientes se produjo un aumento sostenido de la densidad poblacional, debido a la constante entrada de individuos durante el desarrollo vegetativo y a la baja tasa de mortalidad, probablemente por la larga longevidad. Posteriormente, se observan individuos tenerales que han cumplido su estado preadulto dentro del cultivo y van a ser responsables del pico máximo poblacional. Los picos poblacionales se suceden en el período reproductivo del hospedero, en este momento se iguala la proporción de hembras y machos. Generalmente, estos picos son seguidos de descensos bruscos de la población, debido al abandono del campo por la senescencia del cultivo. Este, es realizado en gran cuantía por las hembras, las cuales buscan otros hospederos más aptos que le provean de alimento a su descendencia; por lo tanto, la proporción sexual favorece a los machos, los que no son buenos colonizadores y, por ende, tampoco emigradores. Finalmente, el tiempo de duración del ciclo biológico de estos crisomélidos y la forma de las cunas poblacionales, hacen inferir que solamente se sucede una generación a lo largo del ciclo del cultivo de caraota. CONCLUSIONES Las fluctuaciones poblacionales de las tres especies estuvieron estrechamente relacionadas a la naturaleza temporal del cultivo durante los dos años. La proporción del número total de adultos de A. arcuatus: A. ruficornis. C. equestris quedó representada numéricamente por cifras aproximadas de 6:3: 0,5 durante los dos ciclos del cultivo, habiendo mostrado densidades totales distintas y estando en lugares y fechas completamente diferentes. La interpretación de las fluctuaciones poblacionales de las tres especies durante un ciclo de cultivo de caraota, permite explicar el proceso general siguiente: la colonización del cultivo se realiza principalmente por hembras, cuando las plantas muestran sus primeras hojas. La población va en ascenso sostenido durante el período de desarrollo vegetativo del cultivo, hasta llegar a un pico máximo, que puede llegar a mantenerse unas semanas. En ese momento se detectan individuos tenerales que han cumplido su estado preadulto y van a ser responsables de dicho pico poblacional, la proporción sexual se encuentra equilibrada. Finalmente, ocurre el abandono del campo por la senescencia del cultivo por lo que la proporción sexual favorece a los machos, ya que las hembras han emigrado para otros campos que aseguran la supervivencia a su descendencia. A lo largo de un ciclo de cultivo de caraota, se sucede una solo generación de las especies estudiadas. SUMMARY K,W Andrector. Gynandrobrotica. Populational fluctuation, Phenology, colonization, emigration, Phaseolus. BIBLIOGRAFIA 1. CENTRO INTERNACIONAL DE AGRICULTURA TROPICAL (CIAT). Una escala de desarrollo para el frijol propone el CIAT. Hojas de frijol 5(2):12.1983. 2. DEBROT, C. y C. BENITEZ DE ROJAS. El virus del mosaico del frijol, Vigrw sinensis Endl., (Cowpea Mosaic Virus). Agronomía Trop. 17(5):315.1967. 3. DEBROT, C. y F. CENTENO. Estudios de las enfermedades virales de la soya en Venezuela. In: Jornadas Técnicas CENíAP '84. Centro Nacional de Investigaciones Agrop. Maracay, Ven. 1984. (Resúmenes). p. 4349. Publicación Especial No. 142. 4. FELACO, F. Algunos aspectos sobre colonización de insectos en soya, Glycine max (L.) Merril. Trabajo de Grado. Maracay, Ven. UCV. Fac. Agr.1983. 80 p. 5. FERNANDEZ YEPEZ, F. y C.J. ROSALES. Guía de Entomología Agrícola. Tercera Edición. Maracay, Ven. UCV. Fac. Agr. 1984. 142 p. 6. FORSYTHE, H. Jr. and G. GYRISCO. Determining the appropiate transformation of data fron insect control experiments for use in the analysis of variance. J. Econ. Entomol.54(5):859861. 1961. 7. GUAGLUMI,P. Insetti e aracnidi delle piante comuni del Venezuela segnalati nel perdo 19381963. Relazionie e Monografie Agrarie Subtropicali e Tropicali. 86, Instituto Agronomico per l'Oltremare. Firenze. 1966.391p. 8. GUTIERREZ, V., M. INFANTE y A. PINCHINAT. Situación del cultivo del frijol en America latina. Call, Centro International de Agricultura Tropical (CIAT).1975. 33 p. Serie ESl9. 9 KOGAN, M., W. RUESINK and K. McDOWELL. Spatial and temporal distribution patterus of the Bean leaf Beetle, Cerotoma trifurcata (Forster), on soybeans in lllinois. Environrn. Entomol. 3:607617. 1974. 10. LITTLE, M. and F. HILLS. Métodos estadísticos para la investigación en la agriculture. Trad. A. De Paula Crespo. Mexico, Edit. Trillas. 1984. 270p. 11. MONZON, D., R. MATERAN, P. ANGARITA, A. DAGERT, E. SOTO y J. PACHECO. Apuntes de Estadística '64. Maracay, Ven. UCV. Fac. Agr. 1964. 219 p. (Multigrafiado). 12. SCHOONHOVEN, A. Pests of beans in latin America and their control. In: Pest of grain legumes: ecology and control. New York, Academic Press! p. 151165. 13. SOKAL, R. y F. ROHLF. Biometría: principios y métodos estadisticos en la investigación biológica. Trad. M. Lahoz león. Madrid, Edt. Blume.1969. 832 p. 14. STEEL, and J. TORRIE. Principles and procedures of statistics: with special reference to the biological sciences. New York, Mc Graw Hill. 1960.418 p. 15. VELAZQUEZ, J. y C. CARDONA. Comparación de métodos de mueskeo y desarrollo de un mueskeo secuencial para crisomélidos y saltahojas en frijol. Rev. Colomb. Entomol. 8(34):1118. 1982. 16. WALDBAUER,G. and M.KOGAN. Bean leaf Beetle bionomics and economic role in soybean agroecosystems. In: Proc. World Soybean Research. Conf., lllinois. p. 619628. 1976. 17. YEPEZ GIL, G. Estudio sobre algunos aspectos de la bialogía y ecología de Andrector arcuatus Olivier, A. ruficornis Olivier y Gynandrobrotica equestris Fabricius (Coleoptera: Chrysemelidae). Trabajo de Grado. Maracay, Ven. UCV. Fac. Agron. 1982. 84 p. 18. YEPEZ GIL, G. y A. MONTAGNE. Información sobre métodos de muestreos absolute y relative de A. arcuatw y A. ruficorna (Coleoptera: Chrysomelidae). 8., Congreso Venerolano de Entomología (Resúmenes). Barquisimeto. 1983. 19. YEPEZ GIL, G. y A. MONTAGNE. Estudio sobre algunos aspectos de la biologia de Andrector arcuatw Olivier, A. ruficornis Olivier y Gynandrobrotica equestri Fabricius (Coleoptera: Chrysomelidae), plagas de importancia en carsota. Bol. Ent. Venez. N.S. 4(S):3341. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||