Agronomía Tropical 49(2):187-199. 1999.
DESARROLLO DEL NÍSPERO
DURANTE LA FASE
JUVENIL DEL CRECIMIENTO
CONTINUO
JUVENIL DEL CRECIMIENTO CONTINUO
Trabajo parcialmente financiado por el Concejo de Desarrollo
Científico,
Humanístico y Tecnológico (CDCHT) de la Universidad
Occidental
Lisandro Alvarado (UCLA).
Profesores Lisandro Alvarado. Decanato de Agronomía.
Postgrado de
Horticultura. Apartado 400.
Barquisimeto, estado Lara. Venezuela.
RECIBIDO: noviembre 26,1998.
RESUMEN
Palabras Clave: Manilkara zapota, plántula, fase juvenil, eje epicotilar, ramificación.
ABSTRACT
Key Words: Manilkara zapota, epicotyl axis, continuous growth, juvenile phase, branching.
INTRODUCCIÓN
El níspero Manilkara zapota Royen, llamado en México y países Centroamericanos "Chicozapote", es un árbol perennifolio nativo de América Tropical (Popenoe, 1920), cuya fruta probablemente es la mas apreciada entre las Sapotáceas, familia a la cual pertenece (León, 1968). La fruta es una baya de forma, tamaño y color de la pulpa muy variables (León, 1968; Roth y Lindford, 1972); con cinco o ninguna semilla (Maciel et al., 1996). La germinación es epígea, y la emergencia total puede alcanzar el 70% a los 50 días (Maciel et al., 1996).
El níspero como todas las plantas superiores pasa por las fases de embrión, juvenil y adultez durante su ciclo de vida (Hartmann et al., 1990). El patrón normal del crecimiento del níspero, desde la fase embrionaria hasta alcanzar la adultez, ha sido descrito por Hallé et al. (1978) como perteneciente al modelo de Aubreville. Este modelo se caracteriza por presentar un tronco monopodial de crecimiento rítmico, filotaxia espiralada y ramas en verticilos. Las ramas son plagiotrópicas y de crecimiento modular. Los módulos laterales son de floración lateral y de crecimiento indefinido. Sin embargo, para el níspero no se conocen mayores detalles sobre las etapas iniciales de la vida de la planta, germinación, emergencia, formación del eje ortotrópico e inicio de la ramificación. La fase juvenil, que va desde germinación hasta cuando aparecen las primeras flores (Hackett, 1983), tiene una duración de 6 a 8 años según Popenoe (1920), requiere ser estudiada en cuanto a los cambios morfológicos y fisiológicos de transición hacia la fase adulta.
Algunas especies forman ramas laterales fuertes y vigorosos en la base del tronco o eje principal, mediante el proceso de ramificación por silepsis. Este proceso consiste en la formación de ramas por la entrada en actividad de primordios laterales de manera simultánea con el crecimiento y desarrollo del eje principal (Champagnat, 1954; Maggs, 1960). En el níspero las primeras ramas laterales que se forman, son plagiotrópicas y se originan mediante el proceso de silepsis (Aubréville, 1964; Hallé et al., 1978). Sin embargo, la ramificación siléptica del níspero durante su fase juvenil no ha sido suficientemente estudiada.
En este trabajo se reseña cronológicamente la evolución morfológica de estructuras y órganos que se desarrollan sobre el eje epicotilar, desde el inicio de la germinación hasta cuando el meristemo apical entró por primera vez en reposo, periodo que duró aproximadamente 18 meses.
MATERIALES Y MÉTODOS
La presente investigación se realizó en las instalaciones del Postgrado de Horticultura, Universidad Centro Occidental Lisandro Alvarado, Barquisimeto Venezuela (10º 05' LN; 510 m.s.n.m. de altitud).
Las plántulas se obtuvieron de semillas de frutos maduros adquiridos en plantaciones comerciales de la altiplanicie de Maracaibo, estado Zulia, al noroccidente de Venezuela. Las semillas fueron colocadas en bandejas de 20 cm de profundidad y llenas con un sustrato conformado por la mezcla de arena y fibra de coco en la proporción 1:1 en volumen (Maciel et al., 1996). La germinación y emergencia tuvo lugar bajo condiciones de umbráculo con 80% de restricción de luz, temperatura promedio diaria de 26±4º C y 65±15 % de humedad relativa.
Las plántulas fueron transplantadas a bolsas de polietileno contentivas del mismo sustrato de germinación, cuando se evidenció el inicio de formación de la primera hoja verdadera. De una población superior a 600 plántulas, se seleccionaron 90 por su uniformidad en tamaño, color y número de hojas a los dos y medio meses de edad, desde la emergencia. Estas plántulas presentaban entre una y dos hojas verdaderas, un tamaño de 10 a 12 cm y mantenían sus cotiledones funcionales. En ese momento, se formaron dos lotes de 45 plántulas cada uno. Un lote permaneció bajo el umbráculo usado para germinación y el otro se colocó bajo una estructura abierta con techo transparente de fibra de vidrio. En ambos casos, las plantas fueron regadas, fertilizadas y en general, tratadas de manera similar.
La evolución morfológica de la planta fue puesta en evidencia mediante la observación macroscópica y continuada de la aparición de estructuras y órganos sobre el eje epicotilar durante su ontogenia, siguiendo los conceptos básicos sobre arquitectura de las plantas establecidas por Hallé et al. (1978).El periodo reseñado en este trabajo abarcó desde el inicio de la germinación hasta cuando el meristemo apical del eje epicotilar paralizó su crecimiento por primera vez y se transformó en un yema que entró en reposo. Durante este período, se registró cronológicamente la aparición, forma, número y ubicación de estructuras y órganos sobre el eje epicotilar.
RESULTADOS Y DISCUSIÓN
La plántula al erguirse con motivo de la emergencia, pone al descubierto de abajo hacia arriba, el "collect" o zona de transición hipocotilo - radícula, el hipocotilo, el nudo cotiledonar, los cotiledones y el epicotilo (Figura 1.). El hipocotilo relativamente largo de 5 a 7 cm, inició tempranamente un proceso de suberización mediante el cual, la porción entre la base de la radícula y el nudo cotiledonar se recubrió de suber a lo largo de un período de 10 a 12 meses.
Los cotiledones a partir de la emergencia quedaron perpendiculares al eje epicotilar e iniciaron funciones como órganos fotosintetizantes, contribuyendo tempranamente a independizar a la plántula de sus reservas, característica típica de especies de semillas pequeñas (Ng, 1978). Las hojas cotiledonares de pecíolos muy cortos o casi sésiles, continuaron creciendo durante los primeros cinco meses posteriores a la emergencia, hasta alcanzar un tamaño final de aproximadamente 2,5 veces mayor que el inicial (Tabla 1). Los cotiledones que fueron alargados, elipsoidales y redondeados , presentaron limbos uniformemente planos gruesos y coriáceos, con nervadura central bien definida y ramificaciones laterales que alcanzaron varios órdenes. Los cotiledones completaron su crecimiento cuando ya se habían formado tres a cuatro hojas verdaderas y persistieron por mas de 18 meses sobre la planta. La presencia de éstos se manifestó mediante cambios de color en el limbo, del verde al amarillento y luego al marrón, después de lo cual abcisaron.
|
CUADRO 1. Tamaño de los cotiledones fotosintetizantes desde emergencia hasta máximo tamaño en umbráculo (UMB) y estructura de techo transparente (ETT) n=45. |
||||||||
|
|
||||||||
|
|
PERIODO | |||||||
|
| ||||||||
|
Sept. |
Oct. |
Nov. |
Dic. | |||||
|
|
|
|
| |||||
|
Largo |
Ancho |
Largo |
Ancho |
Largo |
Ancho |
Largo |
Ancho | |
|
|
||||||||
|
ETT |
2,58 |
1,20 |
3,0 |
2,0 |
4,5 |
2,5 |
4,7 |
3,0 |
|
UMB |
2,58 |
1,20 |
3,2 |
1,7 |
4,5 |
2,3 |
4,5 |
3,0 |
|
|
||||||||
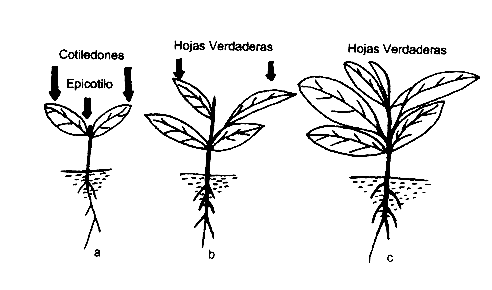 |
|
Figura1. Esquema de la plántula de níspero en tres estados evolutivos: a) Plántula con cotiledones, b) Con dos hojas verdaderas, C) Con cinco hojas verdaderas. |
La primera hoja verdadera se presentó en posición decusada con los cotiledones y quedó completamente expuesta después de 45 días de emergida la plántula. Las hojas que aparecieron de manera espiralada sobre el eje, fueron simples, lanceoladas o elípticas, con ápice acuminado o agudo y de base atenuada o cuneada. La hoja presentó vernación conduplicada, nervadura pinnatinervia y pecíolo de corto a muy corto. El pecíolo ligeramente acanalado adaxialmente presentó un par de estipulas tempranamente caedizas, las cuales se localizaron a nivel de la inserción con el tallo.
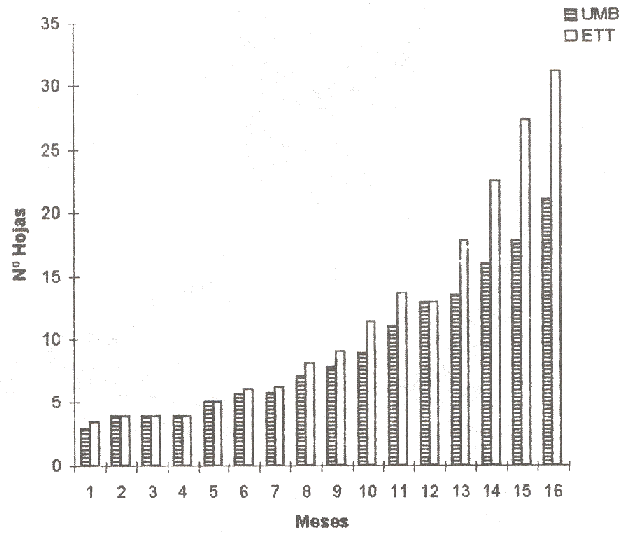
El eje epicotilar se caracterizó por presentar inicialmente una etapa de crecimiento continuo, monopodial, con dirección ortotrópica y filotaxia espiralada. El proceso de crecimiento continuo se mantuvo desde la germinación hasta cuando el meristemo apical se transformó en una yema que entró en reposo (Figura 2). Para el momento de la paralización del crecimiento, la yema apical contiene de 4 a 5 primordios foliares inactivos y está recubierta por al menos tres (3) escamas o prófilos. La paralización del crecimiento ocurrió a lo largo de un período que se extendió de los 15 a los 24 meses de edad. A los 18 meses, las plantas paralizadas alcanzaron una proporción aproximada de 65% en los dos ambientes. Para ese momento, las plantas paralizadas habían formado en promedio de veintidós hojas (22) en el umbráculo y treinta y cinco (35) hojas en la estructura de techo transparente (Figura 3). La transformación en yema terminal del ápice de crecimiento del eje epicotilar , marca la culminación de una etapa de exclusivo crecimiento vegetativo sin cambios transicionales dentro de la fase de juvenilidad que caracteriza esta especie.
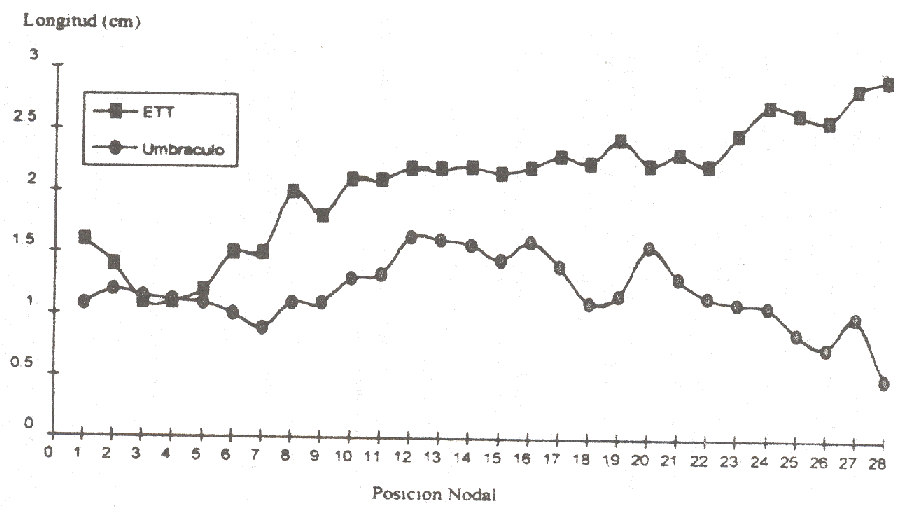
Durante la etapa del crecimiento continuo del eje, los entrenudos inicialmente muy cortos en la zona basal ( 1.0 cm) paulatinamente aumentaron en longitud hasta alcanzar un máximo de 2.0 a 3.0 cm, se mantuvieron allí variando entre pequeños máximos y mínimos y disminuyeron abruptamente cuando el ápice inició la paralización del crecimiento (Figura 4). Desde un comienzo la longitud internodal siempre fue mayor en las plantas bajo ETT que en las de umbráculo, para posiciones nodales similares.
Ramificación
La aparición de ramas se inició a los 9 meses de edad de las plántulas y ocurrió primero en aquellas que ramificaron en las posiciones nodales mas básales. Frecuentemente dos o tres, en menor proporción una a cuatro, ramas aparecieron seguidas o separadas por un nudo sobre la espiral filotáxica a manera de un seudoverticilo. Cada rama presentó el entrenudo basal, el hipopodium muy largo (12 a 15 cm), seguido de dos o tres entrenudos que se acortan abruptamente. Esta rama de posición horizontal, se ramificó simpodialmente cerca del ápice y en el mismo plano, para formar un subsiguiente orden, con dos nuevas ramas en posición dística.
 |
|
Figura 2. Esquema del ápice de crecimiento del eje epicotilar del níspero a) Entrando en reposo, b) En estado de reposo, c) Reiniciando crecimiento. |
 |
|
Figura 3. Número de hojas de plantas en umbráculo (UMB) y estructura de techo transparente (ETT) a los 16 meses de edad en níspero. |
 |
|
Figura 4. Longitud internodal y posición nodal (cm) de plantas en Umbráculo (UMB) estructura de techo transparente (ETT) a los 16 meses de edad desde emergencia en níspero. |
La formación de ramas laterales de manera simultánea al crecimiento del eje epicotilar, constituye un proceso de silepsis el cual parece ser característico de esta especie, es que ya había sido observado y descrito por Aubreville (1964) y Hallé et al. (1978). La aparición de las ramas ocurrió al final de la etapa del crecimiento continuo y antes de la entrada en reposo de la yema terminal. En el umbráculo, las ramas formadas se ubicaron entre las posiciones nodales 18 a la 28; mientras que bajo la ETT, las ramas se localizaron entre los nodos 27 y 42 ( Figura 5). Las ramas adaptaron posiciones aproximadamente perpendiculares al eje, por lo que fueron consideradas como plagiotrópicas.
El eje epicotilar creció continuo e indeterminadamente por un período de hasta 24 meses, después del cual el meristemo apical se transformó en yema. Este eje se caracterizó por presentar tres (3) zonas diferenciables (Figura 5):
- Una basal no ramificable, ubicada entre los cotiledones y las posiciones nodales 18 (umbráculo) y 27 ( ETT).
- La intermedia o de ramificación, donde se insertaron las dos o tres ramas que usualmente se formaron sobre cada eje.
- La distal conformada por una porción de 9 a 11 nudos sin ramas, por debajo de la yema apical en reposo.
Las probabilidades de ubicar las pocas ramas, usualmente consecutivas, sobre una zona de ramificación siempre mayor a 10 nudos así como la época de ocurrencia del fenómeno, resulta muy difícil de predeterminar. Estos procesos pueden ser repuesta a fenómenos endógenos característicos de la especie.
CONCLUSIONES
- La planta mostró una etapa de crecimiento continuo que persistió por 18 o más meses y fue desde la germinación hasta cuando el ápice epicotilar se transformó en una yema en reposo.
- La ramificación se inició a los 8 meses mediante la emisión de dos a tres ramas, por encima del nudo 16 y por debajo del décimo desde el ápice epicotilar.
- El eje epicotilar presentó tres zonas diferenciables, una basal y una apical sin ramificaciones y una intermedia donde aparecieron las ramas.
- La longitud internodal y el tamaño de la planta fue menor en el lote de planta del umbráculo como consecuencia de la restricción de la luz.
- Después de establecido el primer verticilo, el eje epicotolar ha alcanzado un porte y grosor tal que la planta puede ser injertada.
BIBLIOGRAFÍA
Aubreville, A. 1964. Les Sapotaceae. Taxonomie et phytogeographie. Adansonia. Mem. Nº 1. pp. 157.
Hackett, P. 1954. Recherches sur les rameaux anticipés des vegetaux ligeux. Rev. Cytol. Biol. Veg. 15:1-51.
Champagnat, P. 1954. Recherches sur les rameaux anticipès des vegetaux ligneux. Rev. Cytol. Biol. Veg. 15: 1-51.
Hallé, F., R.A. Oldeman and P.B. Tomlinson. 1978. Tropical Trees and forests: an archichectural analysis. Springer Verlag. N.Y. 673p.
Hartmann, H., D. Kester and F.Davies. 1990. Plant propagation. Principles and practices. Printice Hall, Englewood Cliffs. 647p.
León, (J). 1968. Fundamentos botánicos de los cultivos Tropicales. Inst. Interamericano Ciencias Agrícolas. San José. Costa Rica. 487p.
Maciel, N., D. Bautista y J. Aular. 1996. Características morfologicas del fruto y la semilla, y procesos de Germinación y Emergencia del Níspero Manilkara achras (Miller) Fosberg. Proc. Interamer. Soc. Trop. Hortic. 40:188-194.
Ng. E.S.P. 1978. Strategies of establishment in Malayan forest Trees. In: Tropical trees as living systems. P.B. Tomlinson and M.H Zimmermann (Edits)Cambridge Univ. Press. pp: 129-162.
Popenoe, W. 1920. Manual of Tropical and Subtropical Fruits. Mc. Millian. New York (Reprinted by Hajner. Press. New York. 1974. 474 p.
Roth, I. and H. Lindford. 1972. Desarrollo y Anatomia del fruto y de la semilla de Achras sapota L. (Níspero. Acta Botánica Venezuelica 7( 1-4):121-141
Zimmermann, M.H. 1971. Flowering in crabapple seedling methods of shorting the juvenile phase.J. Amer. Soc. Hort. Sci. 96 (4): 404-411.