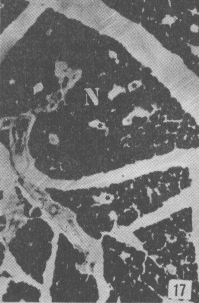|
Veterinaria Tropical 14: 53-83. 1989 APORTES ESTRUCTURALES E HISTOQUIMICOS SOBRE Gisela C. García C. Universidad
Central de Venezuela |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
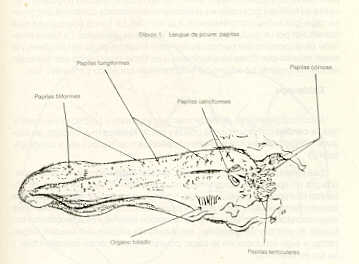
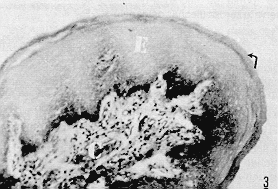
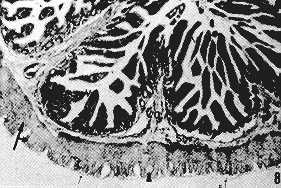
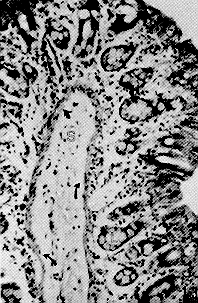
RESUMEN Se describen las características histológicas del tubo digestivo y las glándulas salivales del picure (Dasyprocta rubrata), observadas en cortes coloreadas con Hematoxilina -Eosina, Tricrómico de Gallego y Fontana -Mas- son. Igualmente, fue determinado el tipo de sustancias mucosas presentes en el tracto digestivo mediante las técnicas Ácido Peryódico de Schiff (PAS), Alcian Blue (AB) y la combinación de las técnicas PAS/AP. Se estudia con mayor interés las similitudes estructurales, o la falta de ellas, con otros roedores histricomerfos de la fauna silvestre venezolana, como son el chigüire (Hydrochoerus hydrochaeris) y la lapa (Agouti paca). INTRODUCCIÓN El picure (Dasyprocta rubrata) es una especie de la fauna silvestre venezolana utilizada como alimento en ciertas áreas rurales del país, por lo que puede considerarse como una alternativa proteica potencial que mejoraría la calidad de la dieta de los habitantes de dichas áreas. Además, se reproduce un cautiverio y podría aprovecharse también su piel. La familia Dayproctidae se caracteriza morfológicamente por presentar una longitud cabeza-cuerpo de unos 41,5 a 62,0 cm, con una cola que oscila entre 1,0 y 3,5 cm. Presenta tres de dos con uñas que le sirven de apoyo a las patas posteriores. El pelo es más largo y espeso en la parte posterior del lomo (9). Se localizan desde México hasta el límite de la vegetación peremnemente verde en la Argentina (22). En Venezuela son muy conocidos, especialmente en las regiones conuqueras debido a los daños que causan. El picure es un animal de cacería con hábitos diurnos y nocturnos, que generalmente anda solo (28). Se reproduce todo el año, con 2 o 4 crías por parto (8). En su habitat natural come frutas y raíces (8, 11). No es cavador, pero construye zorreras (28). Es importante aportar conocimientos anatómicos para interpretar integralmente su biología, junto a otros aspectos como su fisiología, reproducción, nutrición y sus posibles patologías. En las literaturas consultadas existen pocas referencias sobre: el picure, especialmente en cuanto al aspecto histológico del tubo digestivo, por lo que este trabajo se basa en la comparación de los aspectos histológicos generales del tubo digestivo de esta especie con los de otros roedores histricomorfos de la fauna silvestre venezolana, como son el chigüire (Hydrochoerus hydrochaeris) y la lapa (Agouit paca). El predominio de acinos mucosos en glándulas salivales de la boca y la faringe, aunado al epitelio nucosentor del estómago, al desarrollo de las células caliciformes en las glándulas de Lieberkühn, y los adenómeros mucosos de las glándulas de Brunner, son observaciones morfológicas que motivaron la realización del estudio histoquímico. MATERIALES Y MÉTODOS Se procesaron muestras de ocho picures provenientes del centro del país. Se eutanaciaron utilizando la combinación Rompun (Xilacina) y Ketalar (Ketamina) con rangos de dosis que oscilaron entre 0,3 y 0,5 mg/kg y 10 y 30 mg/kg, respectivamente. Las secciones obtenidas fueron identificadas y fijadas en formol neutro al 10% y procesadas bajo la técnica convencional de inclusión en parafina. Los cortes obtenidos al microtomo fueron de 5 de espesor. Se colorearon con las técnicas: hematoxilina y eosina (H-E), tricrómica de Gallego y Fontana Masson para el Procesamiento histológico (16). Para el análisis histoquímico se realizaron las técnicas: ácido peryódico de Schiff (P.A.S.), Alcian Blue (AB) a pH 1,0 y 2,5 y la combinación de las técnicas PAS/AB, para los estudios cualitativos de mucosustancias (16, 27). Las fotografías se tomaron en un fotomicroscopio Zeiss con películas Kodak y Agfa Asa 64 y 100 con los objetivos 2,5 X; 6,3 X; 10 X y 25 X. RESULTADOS Resultados Histológicos Lengua Los cortes histológicos del órgano evidencian un revestimiento epitelial plano estratificado queratinzado. Este descansa sobre un corion de tejido conjuntivo que varía de moderadamente de denso a laxo. En plano más profundo se dispone la musculatura del órgano, constituida por fibras musculares estriadas dirigidas en varios sentidos, (longitudinal, transversal, oblicuo). Entre los haces musculares se pueden observar acúmulos de Células adiposas. En la región basal de la lengua se describen los acinos mucosos y serosos de las glándulas salivales linguales (Figuras 1 y 5). La región dorsal de la lengua posee un corion papilífero que forma papilas macroscópicas filiformes , distribuidas desde la punta del órgano hasta las cercanías de su base, donde son sustituidas caudalmente por papilas cónicas y lenticulares (Figura 3). En todas se destaca el grosor de la capa cornea . Las papilas filiformes están acompañadas en su distribución por las papilas fungiformes, de número notablemente menor y capa córnea menos evidente. Estas papilas adquieren un desarrollo más relevante en la base del órgano y alrededor de las papilas caliciformes, las cuales a su vez, en número de dos (Dibujo 1), se disponen formando un ángulo con vértice dirigido caudalmente. Cabe destacar que la capa córnea es más exigua en estas papilas caliciformes. El picure posee órgano foleado o papilas foliáceas ubicadas a nivel postero-lateral (Dibujo 1 ). En sus surcos interpapilares desembocan las glándulas de Von Ebner, tal cual se observan en las papilas caliciformes (Figura 1 ). Se evidencian botones gustativos en las papilas caliciformes, fungiformes y órgano foliado.
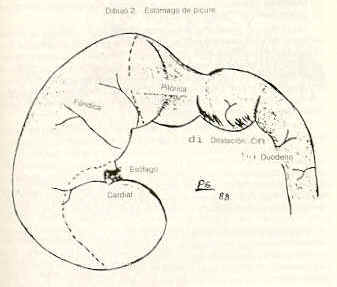
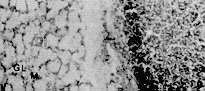
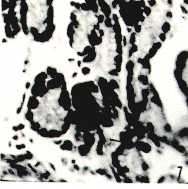
Glándulas salivales mayores Las glándulas parótidas presentan una cubierta conjuntiva que las divide en lóbulos y lobulillos. Poseen ganglios linfáticos en el conjuntivo interlobulillar. Están compuesta por acinos mucosos puros (Figura 2). Las glándulas mandibulares están organizadas en lóbulos y lobulillos. Sus acinos son mucosos puros (Figura 4). Las glándulas sublinguales presentan características similares a las descritas en las glándulas antes citadas. Esófago Se hizo su estudio tomando muestras de tres áreas: trayecto cervical (tercio craneal), trayecto torácico (tercio medio) y trayecto abdominal (tercio caudal o distal). En los cortes histológicos estudiados se observó el revestimiento querantizado del órgano que descansa sobre un corion aglandular. La muscular de la mucosa está formada por fibras musculares lisas; es poco evidente en la parte cervical, adquiere mayor desarrollo en la parte torácica (Figura 6) y alcanza su máxima expresión en el tercio posterior donde conforma una gruesa capa que rodea completamente a la mucosa. La túnica submucosa está constituida por un tejido conjuntivo laxo; carece de glándulas. La túnica muscular se dispone en dos planos: el interno de fibras musculares circulares y el plano externo con fibras longitudinales. Las fibras musculares son estriadas en todo el órgano. La adventicia está formada por tejido conjuntivo laxo. Estómago Macroscópicamente se evidencian tres regiones (r.) o partes bien definidas: la cardial con una mucosa pálida y lisa; la fúndica muy irrigada, de pliegues longitudinales y transversales; la pilórica con mucosa lisa y pálida (Dibujo 2). Los cortes histológicos de la región cardial muestran una mucosa revestida por un epitelio cilíndrico simple mucosecretor. A nivel del corion se observan dos tipos de glándulas; unas, tubulosas simples ramificadas, de luz y corion interglandular moderadamente amplios, con células exclusivamente mucosecretoras, justo en la zona de transición epitelial y de adenómeros tortuosos. El otro tipo del glándulas, son tubulosas simples ramificadas, largas, rectas y algo tortuosas en la base; poseen escaso corion interglandular y su luz es estrecha. Los elementos celulares que la conforman son: células mucosas, células parietales abundantes, células principales y enteroendocrinas (Figura 7). La muscular de la mucosa es muy desarrollada, principalmente en la zona de transición epitelial; emite prolongaciones interglandulares. La región fúndica está revestida por un epitelio cilíndrico simple mucosecretor. Se torna más bajo a nivel del cuello (células mucosas). Al profundizar en la glándula, las células epiteliales son sustituidas por células de mayor tamaño, con núcleo central y citoplasma eosinofílico (células parietales), muy abundantes en el cuerpo glandular, en comparación con el fondo de las mismas. También se localizan células más pequeñas que las anteriores, de núcleo basal y citoplasma basófilo (células principales), las cuales predominan en el fondo glandular. Con la reacción Fontana-Masson, se evidencian las células enteroendocrinas con sus características granulaciones citoplasmáticas oscuras. Las glándulas de esta región son más cortas que las de la región cardial, pero el revestimiento epitelial mucosecretor es más prominente. El tejido conjuntivo interglandular, aunque escaso, es más abundante al compararlo con el de la región antes mencionada, resultando moderadamente abundante en la zona sublandular. La muscular de la mucosa es menos desarrollada al compararla con la de la región cardial, también emite prolongaciones interglandulares.
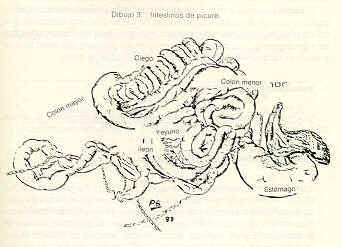
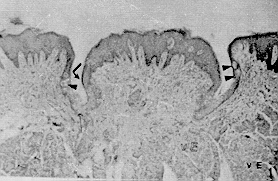
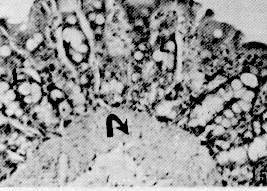
La región pilórica revela una mucosa de aspecto foliado, revestida por un epitelio cilíndrico mucosecretor (Figura 9). Las glándulas son del tipo tubulosas. Las células parietales y principales de la región fúndica son sustituidas por células de citoplasma eosinofílico. Se evidencian células enteroendocrinas en esta región. La mucosa es más amplia que la de las regiones antes descritas. El corion interglandular es abundante. La muscular de la mucosa es más desarrollada que en la región fúndica. El estómago presenta las otras túnicas típicas del tubo digestivo. La submucosa es muy vascularizada, especialmente en la región fúndica, está formada por tejido conjuntivo laxo; carece de glándulas. La túnica muscular es de fibras lisas dispuestas de manera similar a la descrita en el esófago. Sin embargo, en la región pilórica es posible observar áreas con un tercer plano de fibras oblicuas y circulares. El órgano está recubierto por una serosa típica. Intestino delgado El intestino delgado del picure puede dividirse en tres regiones o áreas: duodeno, yeyuno e ileon. Duodeno. En su parte inicial presenta una dilatación que luego disminuye su calibre hasta uniformizarlo (Dibujo 2). Las secciones histológicas muestran características similares en todo el duodeno. La mucosa es plegada, principalmente a nivel de la dilatación. Presenta vellosidades intestinales y pliegues permanentes. El epitelio es cilíndrico simple con chapa estriada y células caliciformes. A nivel de corion se observan las glándulas de Lieberkühn, con células de citoplasma claro alternándose con la presencia de células cilíndricas de citoplasma basófilo, y con células piramidales de citoplasma débilmente eosinofílico y núcleo basal localizadas en el fondo de la glándula. La muscular de la mucosa está presente, siendo interrumpida de trecho en trecho por los acinos y/o conductos excretores de las glándulas de Brunner. La submucosa, de tejido conjuntivo laxo, es ocupada principalmente por las glándulas de Brunner (Figura 11 ). Las células de estas glándulas son cilíndricas mucosecretoras, con núcleo basal. Pueden extenderse hasta las primeras porciones del yeyuno. Las placas de Peyer se ubican en la submucosa, pudiéndose extender hasta el corion. Las túnicas muscular y serosa presentan caracteres similares a los de los órganos anteriormente descritos. El yeyuno presenta un epitelio similar al del duodeno. Las vellosidades intestinales son largas; el número de válvulas conniventes (Figuras 10 y 12) es comparativamente mayor respecto al duodeno, se localizan a nivel de borde mesentérico. En general carece de glándulas de Brunner, excepto en su porción inicial donde algunos cortes evidenciaron su presencia. También se puede observar en algunos cortes la presencia de placas de Preyer. Las túnicas muscular y serosa no muestran caracteres diferenciales con las de los órganos precedentes. El ultimo segmento del intestino delgado, el ileon, presenta características histológicas semejantes a las descritas en el yeyuno (figura 8). Intestino grueso Dividido en las siguientes partes: ciego, colon recto y canal anal. el ciego, muy voluminoso; el colon con la parte inicial relativamente gruesa, que se adelgaza gradualmente y conforman asas en forma espira (Dibujo3). Por ultimo se observa el recto y el conducto anal.
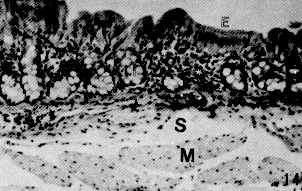
El ciego presenta la mucosa plegada, baja, con glándulas de Lieberkühn tubulosas cortas, separadas unas de otras por abundante tejido conjuntivo laxo, que puede presentar infiltración linfocitaria (Figura 14). El epitelio superficial es cilíndrico simple con chapa estriada, se hace más bajo a nivel de las criptas, siendo sustituido en el fondo de las mismas por células mucosecretoras, evidenciándose células enteroendocrinas por la técnica de Fontan-Masson. La muscular de la mucosa se reduce a una delgada banda que separa el corion de la submucosa. Esta última está formada por tejido conjuntivo laxo (Figura 13). La túnica muscular se dispone en dos planos, con áreas de diferentes grados de desarrollo. La última túnica es una serosa típica. El colón, tal cual se observa macroscópicamente, presenta una mucosa plegada en toda su extensión. El epitelio es similar al descrito en el ciego, pero con glándulas de Lieberkühn más rectas, largas, numerosas y muy próximas entre sí, separadas por escaso corion interglandular, el cual varía de laxo a moderadamente denso (Figura 15). En algunas muestras se observa infiltrado linfocitario. Los elementos predominantes son células mucosecretoras. Se evidenció la presencia de células enteroendocrinas. La muscular de la mucosa es muy desarrollada; alcanza su máxima expresión a nivel del ápice de los pliegues, y la menor en las zonas no plegadas. La sub mucosa está formada por tejido conjuntivo laxo; puede presentar infiltración adiposa. La muscular posee un desarrollo marcadamente mayor que la del ciego, con disposición y naturaleza de sus fibras similares a las descritas en el intestino delgado. La serosa es típica (Figura 16). El recto presenta la mucosa con pliegues menos desarrollados al compararlos con los del colon. Las glándulas de Lieberkühn son más cortas y el corion interglandular más abundante; la muscular de la mucosa se adelgaza en este segmento, siendo más manifiesto en los pliegues. Las otras túnicas presentan características similares a las descritas en el colon. Resultados Histoquímicos Las muestras procesadas y coloreadas con las técnicas P.A.S., Alcian Blue a pH 1,0 y 2,5 y PAS/ AB demostraron los siguientes resultados: Glándulas salivales de boca y faringe. Los cortes coloreados con el reactivo del Acido Peryódico de Schiff (P.A.S.) evidenciaron de moderada a fuerte positividad en todas las glándulas salivales mayores y menores de la boca y faringe, lo que demuestra la presencia de mucosustancias neutras. A nivel de glándulas serosas de la boca (Von Ebner) y faringe (acinos mixtos) también se obtuvo positividad moderada a estas mucosustancias (Cuadros 1 y 2 y Figuras 19, 22 y 25). En el Cuadro 1 se presenta el resumen de los resultados histoquímicos de la boca y la faringe. En referencia a las mucosustancias ácidas, éstas se hacen presentes en las glándulas mucosas linguales y faríngeas, evidenciadas por su fuerte positividad a las reacciones Alcian Blue y PAS/AB efectuadas a pH 1,0 y 2,5 (Figuras 19, 22, 23, 25). En cuanto a las glándulas salivales mayores, se destaca la presencia de estas sustancias, en las glándulas sublinguales, en las cuales la reacción mostró fuerte positividad (Figura 18 y 24) las secciones coloreadas de las glándulas parótidas, demostraron la ausencia de estas sustancias (negatividad) o escasa presencia de las mismas (positividad débil) a pH 2,5 (Figura 17 y 20 ), mientras que las glándulas mandibulares resultaron negativas al AB, lo que manifiesta la ausencia de estas mucosustancias. Estómago Los resultados obtenidos en este órgano demostraron escasa presencia mucosustancias ácidas a nivel del epitelio superficial, dado la negatividad o trazas con la reacción al AB a pH 2,5, mientras que al pH 1,0 los resultados oscilaron entre trazas y positivo débil. Al realizar la reacción del PAS, se demostró la presencia de mucosustancias neutras en todo el epitelio con la respuesta fuertemente positiva.
En relación con las glándulas cardiales, conformadas exclusivamente por células mucosas, se evidenció la presencia de mucinas neutras y sialomucinas, pero ausencia de sulfomucinas (negatividad al AB pH 1,0). Las otras glándulas presentes en la región cardial, al igual que las glándulas de la región fúndica, poseen niveles decrecientes de mucinas ácidas desde la región cardial (Figura 24) hacia la fúndica, con resultados que van de positivo débil a trazas, llegando inclusive a negativa en sentido caudal. Las mucinas neutras se hacen presentes en toda la región cardial, pero disminuyen gradualmente hacia la región fúndica en la medida que las células mucosas son sustituidas por células principales y parietales (figura 26), tornándose más evidente en la región fúndica, donde. los elementos celulares predominantes son las células parietales, principales y enteroendocrinas, en comparación con las células mucosas. En referencia a la región pilórica, se destaca la presencia de mucinas neutras en forma creciente desde la proximidad a la región fúndica hacia el duodeno, haciéndose más manifiesta en la medida en que 1as células mucosecretoras sustituyen a las parietales y principales y llegan a conformar la totalidad de la glándula (Figura 27). La reacción en este caso es fuertemente positiva. En cuanto a las mucosustancias ácidas, se observa su presencia de trazas a positividad débil en el tercio superior, y de moderado a positivo fuerte en los dos tercios inferiores. Similar al caso de las mucinas neutras, las mucosustancias ácidas incrementan su reacción en la medida que se progresa en sentido caudal (Cuadros 3 y 4 y Figura 28).
Intestino delgado. En esta parte del tubo digestivo hay un incremento marcado en la presencia de mucosustancias ácidas, en relación con la producción de las mismas en estómago. Se evidencian en células caliciformes, chapa estriada, glándulas (criptas) de Lieberkühn y glándulas de Brunner, con reacciones que van de positivo moderado a fuerte. También es muy manifiesta la presencia de mucosustancias neutras, que se observan en forma moderada a nivel de chapa estriada y células caliciformes (vellosidades y glándulas de Lieberkühn), y en forma fuertemente positiva a nivel de glándulas de Brunner, tal cual se desprende de los resultados obtenidos en las reacciones de PAS, AB y PAS/AB, cuyo resumen se presenta en los Cuadros 5 y 6.(Figuras 29, 30 y 33). Intestino grueso En esta porción del tubo digestivo se manifiesta también un incremento en la cantidad de mucosustancias ácidas, especialmente del tipo sulfomucinas, ésto en la medida que se progresa en sentido caudal. En el ciego se evidencia la presencia de mucosustancias neutras, en proporcción tal que a pesar de la existencia de mucosustancias ácidas, las neutras casi enmascaran a las mismas (reacción positiva de moderada a fuerte referidas en el Cuadro 8, como otras mucosustancias), como se observa al realizar la reacción PAS/AB (Cuadro 7). Es notable la presencia de estas mucinas neutras, principalmente localizadas en el tercio superior de las glándulas del colon, región donde es más manifiesto el incremento de las mucosustancias ácidas, cuando se comparan las reacciones en el colon espiral (menor) con las reacciones del colon mayor. En cuanto al recto, se observa la presencia tanto de mucosustancias neutras como ácidas en toda la extensión de la glándula de Lieberkühn (Figuras 31 y 32). DISCUSIÓN Y CONCLUSIONES Aspectos Histológicos Las características histológicas de la lengua concuerdan con las características de otras especies, tal cual lo describen (23) y (4). La diferencia macroscópica resaltante, al compararla con la lengua de lapa, estriba en la disposición y número de papilas caliciformes, ya que mientras la lapa posee cuatros papilas dispuestas en dos filas, el picure solo presenta dos papilas, pero dispuestas de manera tal que recuerdan la disposición en V de las papilas caliciformes del hombre (7).
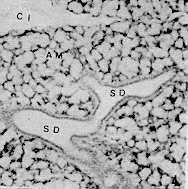
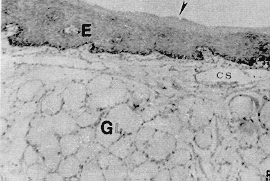
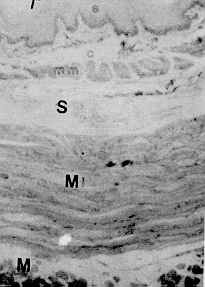
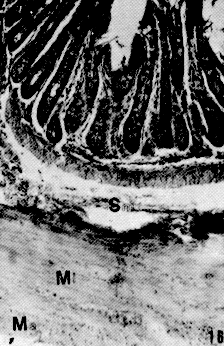
La conformación estructural de las glándulas salivales concuerda con la descrita en otros mamíferos, sin embargo, resalta la constitución de adenómeros mucosos puros en todas las glándulas salivales mayores y linguales excepto en las últimas que estén relacionadas con las papilas gustativas (glándulas serosas de von Ebner); así como glándulas de la faringe donde se pueden encontrar también algunos acinos mixtos. Estas características son comunes en lapa y picure, quizás debido a la alimentación rica en fibras que obliga a producir moco abundante para favorecer el progreso de la ingesta. El esófago posee epitelio plano estratificado querantinizado, similar al de rumiantes y roedores como lapa y chigüire (4, 10, 12, 15, 23), para protegerse del paso de los alimentos. La submucosa carece de glándulas, similar a lo observado en las especies antes mencionadas, excepto en camellos (12); ello se explica por la presencia de queratina en el epitelio del órgano y por la secreción mucosa de las glándulas presentes en boca y faringe. La observación de los cortes del estómago evidencian la ausencia de región (parte) glandular (esofágica) del órgano del picure, esto concuerda con lo citado por (23) y (4) en el sentido que esta región no existe o está muy reducida en carnívoros y hombre, y al igual que en la lapa (10). El estómago macroscópicamente presenta características comunes con el estómago del ratón, donde se observa la parte anterior del estómago con pared pálida y lisa. El cuerpo con pared oscura y corrugada que incluye vasos sanguíneos, y el antro nilórico menos vascularizado y pared también lisa (16). Características similares a las descritas se observaron en la lapa (10). Las secciones histológicas de la región cardial muestran dos tipos de glándulas. Uno formado por células mucosas, similares a las glándulas encontradas en la transición esófago-cárdias de la lapa (10); y en la región cardial del ratón (16) y chigüire (15). El otro tipo de glándula es semejante al encontrado en ratones (16) y la lapa (10), con células mucosas, parietales y principales. A nivel de la región fúndica, llama la atención el gran número de células parietales presentes, acorde con lo reportado en chigüire (15) y lapa (10). La región pilórica presenta características semejantes a las de los otros mamíferos (4, 10, 15, 23, 25). En el intestino delgado es relevante la presencia de una dilatación al inicio del duodeno. Ello es similar a lo observado en otras especies como manatí, lapa y ballenas (10, 20, 25). Las vellosidades intestinales son largas y numerosas y los pliegues permanentes (válvulas comniventes) son muy numerosos y desarrollados, similar a lo observado en lapa (10) y en el chigüire (15). Se pueden encontrar placas de Peyer desde duodeno hasta ileon, acorde con lo reportado en perros (26). El intestino grueso posee un ciego voluminoso como se observa en lapas (10) y equinos (23). El colon, de moderado desarrollo, se adelgaza gradualmente y conforma asas en espiral que son menos manifiestas en comparación con las asas espirales de la lapa, de los suinos y los rumiantes (4, 10). Aspectos Histoquímicos Los resultados obtenidos en la parótida del picure mostraron la presencia de mucinas neutras abundantes y escasas sialomucinas (mucinas ácidas); resultados similares han sido señalados en ratas (3). La glándula sublingual mostró presencia de mucinas neutras y ácidas, tal como se observa en ratas y lapas (3, 10). La glándula mandibular sólo posee mucosustancias neutras, a diferencia de lo observado en ratas y liebres, que además de éstas, poseen mucinas ácidas (3, 18). Las glándulas salivales menores de la boca y la faringe presentan mucosustancias neutras y ácidas en los acinos mucosos, mientras que los serosos sólo demuestran presencia de mucinas neutras; resultados parecidos han sido citados en lapas (10). En el estómago se destaca, a nivel del epitelio superficial, la presencia de abundantes mucosustancias neutras y escasas mucosustancias ácidas, semejante a lo reseñado en peces, anfibios, reptiles, aves y mamíferos (ballenas y lapas) (10, 24, 25). Los resultados obtenidos en las glándulas del estómago evidencian la presencia de mucinas neutras y ácidas, siendo estas últimas más notables en las regiones cardial y pilórica; resultados similares han sido encontrados en peces, anfibios, reptiles, cetáceos y roedores (5, 10, 24, 25). En concordancia con lo observado en peces, anfibios y aves, la chapa estriada del intestino delgado del picure presenta mucinas neutras (24). El hallazgo de mucinas ácidas también concuerda con lo evidenciado en aves, reptiles y ballenas (24, 25), y con lo citado por Fawcett (6) en relación con la presencia de ácido siálico a nivel del gricocálix. En las glándulas de Lieberkühn, se encontraron mucinas ácidas y neutras, lo que se corresponde con lo señalado en aves, peces, reptiles y mamíferos (10, 20, 24, 25). En las glándulas de Brunner se evidencian mucosustancias neutras y ácidas, en reacciones positivas que varían entre moderadas y fuertes, semejante a las lapas, zarigüeyas y manatíes (10, 13, 14, 20, 25), sin embargo, otro trabajo sobre lapas sólo informa sobre mucosustancias neutras en este roedor, pero no de sulfatadas (19); quizás ellos sea debido a las técnicas histoquímicas utilizadas en él. Los resultados obtenidos en el intestino grueso, demuestran que la presencia de mucinas neutras es resaltante en la parte superior de las glándulas de Lieberkühn, mientras que las mucosustancias ácidas predominan en los dos tercios inferiores de ciego y colon. Estos resultados son similares a los observados en bovinos, ratas, ratones y cobayos (1, 2, 21 ). El resultado del recto, donde se observó la presencia de mucinas ácidas y neutras, es también semejante a lo reportado en bovinos y lapas (10, 2). De lo anterior se desprende que la distribución y tipificación de mucosustancias en el picure es muy similar a la observada en otras especies, salvo pequeñas variaciones. SUMMARY In this research are described some histological and histochemical details of digestive tract and salivary glands of the picure (Dasyprocta rubrata), by routine stains such as hematoxiline and eosine (H & E), Gallego's trichrome Fontana-Masson, Periodic Acid Shiff (PAS) and PAS/AB. An attempt was made to compare structural similarities between this rodent, and other hystricomorphus rodent of the Venezuelan wild life, such as capybara (Hydrochoerus hydrochaeris) and lapa (Agouti paca). Figura 1. Papila caliciforme de picure. Se observa el surco (flecha curva) en el cual desembocan los cond. excret. (flecha delgada) de las glándulas de von Ebner (VE). Nótese los botones gustativos (punta de flecha). H & E. 2,5X Figura 2. Glándula parótida. Se aprecian los acinos mucosos (GI. M), y parte del ganglio linfático, en forma de cuña; localizado entre sus lóbulos. H & E. 2,5X. Figura 3. Papila lenticular. Se observa el epitelio plano estratificado (E) con capa córnea (flecha) que descansa sobre el corion (C). H & E. 10X. Figura 4. Glándula mandibular. Nótese la naturaleza mucosa de los acinos (AM), el sistema ductal (SD) y el corion interlobulillar (CI). H & E. 2,5X. Figura 5, Base de la lengua de picure. Obsérvese el epitelio plano estratificado (E) queratinizado (punta de flecha). En el corion destacan capilares sanguíneos (CS) y acinos mucosos de las glándulas linguales. H & E. 10X. Figura 6. Esófago de picure (región torácica). Se observa el epitelio plano estratificado (E) queratinizado (flecha); el corion (C) aglandular y la muscular de la mucosa (m m). Luego se observa la submucosa (S) también aglandular; la muscular del órgano con la lámina interna de fibras circulares (Mi), y la lámina , externa de fibras longitudinales (Me), Retículo de Gridley. 6.3X.
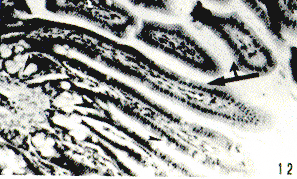
Figura 7. Región cardial de picure. Las flechas señalan las células del sistema enteroendocrino, con las características granulaciones oscuras. Fontana-Masson. 25X. Figura 8. Ileon de picure. Se destaca el gran número de pliegues permanentes o válvulas conniventes (YC), las vellosidades intestinales (Y), la sub mucosa (Sm); los dos planos musculares (flecha doble), en medio de los cuales se observa el plexus nervioso mientérico (punta de flecha). Por último se observa la serosa (flechas delgadas). H & E. 2,5X. Figura 9. Región pilórica. Nótese el aspecto foliado de la mucosa, el epitelio mucosecretor, al igual que el de las glándulas. (Flechas) PAS. 10X. Figura 10. Yeyuno. Detalle de un pliegue permanente donde se destacan las glándulas de Lieberkühn (GI.L), las células caliciformes (Cc) y la submucosa (Sm) H & E. 10X. Figura 11. Duodeno. Corte a nivel de pliegue permanente donde se observa las glándulas de Lieberkühn (gl. L),la muscular de la mucosa (mm) y los acinos mucosos de las glándulas de Brunner (GI-B), localizadas entre el corion y la sub mucosa (Sm). H & E. 10X. Figura 12. Yeyuno. Nótese lo largo de las vellosidades intestinales (flecha doble), las células caliciformes (puntas de flechas finas) en vellosidades y en las glándulas de Lieberkühn (punta de flecha gruesa). Obsérvese el músculo de la vellosidad (-). H & E QX.
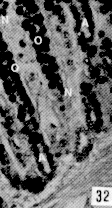
Figura 13. Ciego de picure. Se observa el epitelio cilíndrico simple (E); las glándulas de Lieberkühn, cortas y poco numerosas (L);1a muscular de la mucosa muy delgada (flecha) y la submucosa (S). H & E. 10X. Figura 14. Ciego de picure. Nótese el epitelio (E), las glándulas de Lieberkühn (U) y el infiltrado linfocitario a nivel del corion (*); la delgada banda correspondiente a la muscular de 1a mucosa (punta de flecha); la submucosa (S) y la túnica muscular (M). H & E. 10X. Figura 15. Colon espiral. Se observa el mayor desarrollo de las glándulas de Lieberkühn (L) y de la muscular de la mucosa (flecha). H & E.10X. Figura 16. Colon mayor. Se observan las glándulas de Lieberkühn (L) muy largas y numerosas; 1a muscular de la mucosa (mm) dis puesta en dos planos (interno: circular y externo: longitudinal); la submucosa (Sm); la túnica muscular con sus dos planos: circular (Mi) y longitudinal (ME); por, ultimo se observa la serosa (flecha). Tricrómico de Gallego. 6,3X.
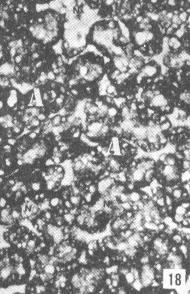
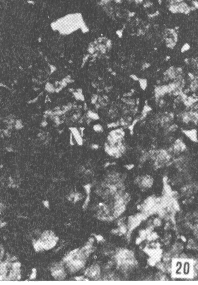
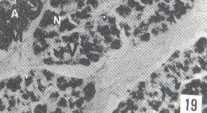
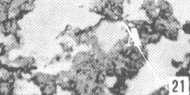
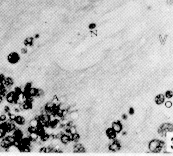
Figura 17. Parótida de picure. Nótese el predominio de mucosustancias neutras (N) en los acinos. Se observan también los conductos excretores (*), un vaso sanguíneo (V) yel tejido conjuntivo que lo tabica (TC). PAS/ AB 2,5X. Figura 18. Glándula sublingual. En el corte se evidencia el predominio de mucosustancias ácidas (A), respecto a la cantidad de mu- cosustancias neutras (N). PAS/AB 2,5X. Figura 19. Lengua. Se observa la presencia de mucosustancias neutras (N) a nivel de glándulas de von Ebner (VE), mientras que las linguales mucosas son positivas tanto a mucosustancias ácidas como a neutras. PAS/AB.10X. Figura 20. Parótida. Nótese la proporción de mucosustancias neutras (N) con respecto a la proporción de las ácidas (A). PAS/AB. 10X. Figura 21. Glándula sublingual. Se observa la reacción positiva a mucosustancias ácidas sulfatadas (A). AB pH = 1,0 2,5X.
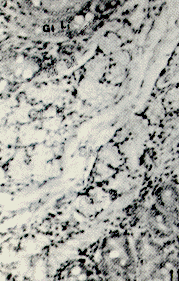
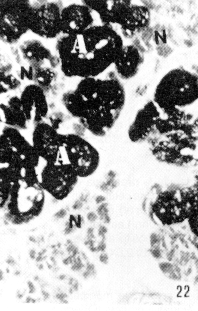
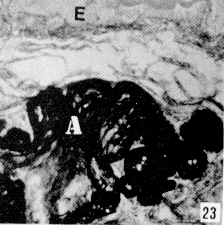
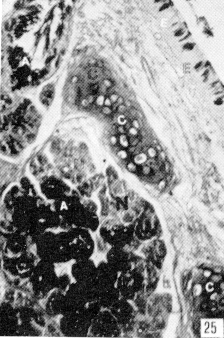
Figura 22. Faringe de picure. Se observa la presencia de mucosustancias neutras (N) en los acinos serosos, y mucosustancias ácidas en los acinos mucosos (A). PAS/AB. 10X. Figura 23. Faringe de picure. En la región orofaringea se observa el epitelio plano estratificado (E) queratinizado y los acinos mucosos positivos a mucosustancias ácidas (A). PAS/AB. 10X. Figura 24. Región cardial. Nótese el epitelio plano estratificado (Ep) alrededor del cual se observan las glándulas positivas tanto a mucosustancias ácidas. como a neutras .(flecha doble) mientras que en el resto del órgano, se observa el epitelio superficial cilíndrico mucosecretor, positivo a sustancias neutras (S), las cuales predominan en las glándulas del resto del órgano (punta de flecha). PAS/AB. 2,5X. Figura 25. Faringe. A nivel de la nasofaringe se : observa el epitelio pseudoestralificado cilíndrico (E) con células caliciformes, y acinos mucosos de secreción ácida (A) y serosos de secreción neutra (N). Nótese la presencia de cartílago hialino (C). PAS/ AB. 10X. Figura 26. Región fúndica. Se evidencia la presencia de mucinas neútras en la superficie epitelial (flecha) y en el cuello de la glándula. PAS/AB. 2,5X.
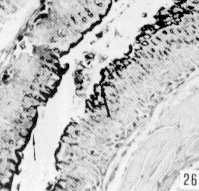
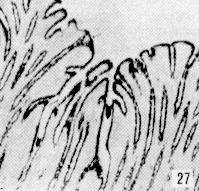
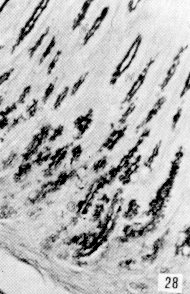
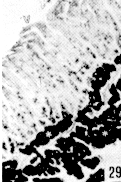
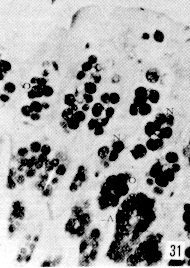
Figura 27. Región pilórica de picure. Se evidencia la presencia de mucinas neutras, tanto en superficie epitelial como en glándulas (fle- chas). PAS. 2,5X. Figura 28. Región pilórica de picure. Se observa el predominio de mucinas neutras en el tercio superior de la glándula (N), mientras que en la base se evidencian, tanto neutras como ácidas (O). PAS/AB. 10X. Figura 29. Duodeno. Se evidencia la presencia de mucinas ácidas, tanto en las células caliciformes de las vellosidades (V), como de las glándulas de Lieberkühn. Las glándulas de Brunner muestran positividad fuerte a mucinas ácidas (*). AB. 2,5X. Figura 30. Ileon. Nótese la presencia de mutinas neutras (N) y de ácidas y neutras (O) en vellosidades. En las glándulas de Lieberkühn se observan tanto neutras (N), como ácidas (A). PAS/AB. 10X. Figura 31. Colon. Se evidencia el predominio de mucinas ácidas (A) en el fondo, mientras que hacia la superficie, lo hacen las neutras (N) y las ácidas y neutras (O). PAS/AB. 10X. Figura 32. Colon. Nótese el predominio de mucinas ácidas en el fondo glandular (A), ácidas y neutras (O) en la parte media y en menor proporción las (N). PAS/AB. 10X. Figura 33. Duodeno. Se evidencia el predominio de mucinas ácidas (A) en glándulas de Brunner Lieberkühn. Se observa la presencia de mucinas neutras tanto en vellosidades como en glándulas de Lieberkühn (flecha corta) y de mucinas ácidas y neutras en vellosidades (cabeza de flecha) PAS/AB. 10X.
BIBLIOGRAFÍA 1. ALTAMANN, G. G. Morphological observations on mucous secretin nongoblet cells in deep crypts of rat ascending colon. Am. J. Anat. 167: 95-117. 1983. 2. ASARI, M. et al. Developmental changes in the Inner Surface structure of the bovine large intestine. Acta Anat. 127: 137-141. 1986. 3. ASOJO, T. A. y AIRE,T. Microsterological and histochemical studies of salivary glands of the giant rat (Cricetomys gambianus, watherhouse). Acta Anat. 117:65-72. 1983. 4. BANKS, WILLIAM. Applied veterinary histology. Baltimore. Edit. Williams & Wilkins. pp. 373-423. 1981. 5. EL -ALFY, M., LEBLOND, C. P. and LEE, E. R. Dinamic histologicof the antral epithelium in the mouse stomach: V. proliferation gradient from the gland base to the isthmus at various times of day. Am.J. Anat. 178: 65-71. 1987. 6. FAWCETT, DON. The cell. Second Edition. Philadelphia. W. B. Saun. ders Company. pp. 35-43. 1981. 7. FAWCETT, DON. Tratado de Histología. 11a. Ed, Trad. Gonzalo Herranz Rodríguez. Madrid, España. Interamericana Mc. Graw-Híll pp. 583-735. 1987. 8. FERNANDEZ YEPEZ, A. y ULLOA, G. Mamíferos de Venezuela. Maracay. U.C.V. Facultad deAgronomía. 102 p. 1969. 9. FERNANDEZ YEPEZ, A. y ULLOA, G. Claves para la determinación de órdenes, familias y géneros de mamíferos en Venezuera. Maracay. U.C.V. Facultad de Agronomía. 65 p. 1970. 10. GARCIA C., GISELA C. Aspectos morfológicos e histoquímicos del tubo digestivode la Lapa (Agouti paca). Trabajo de Ascenso a la categoríade Asistente. Maracay, Venezuela. U.C.V. 80 p. 1984. 11. GONZALEz, Y. de W. Aporte prelimínar sobre hábitos alimenticios y comportamiento durante la alimentación en pícures (Dasyprocta aguti). Resumen. 32 Convención Anual Asovac. Caracas,1982.ln: Acta Científica Venezolana. Vol, 33: 414: Suplemento No, 1. 1982. 12. JAMDAR, M. N. and EMA, A. N. The submucosal glands and orientation of musculature in the Oesophagus of thecamel. J. Anat. 135: 165-171. 1982. 13. KRAUSE, W. J. and LEESON, R. 1969. Studies of Brunner's glands in the opossum. Am. J. Anat. 126: 255-274. 1969. 14. KRAUSE, W. J. and CUTTS, J. H. Morphological observations on the papillae of the opossum tongue. Acta Anat. 113: 159-168. 1982. 15. LEAL M., L. J. Contribución al estudio histológico del tubo digestivo del Hidrochoerus hidrochaeris Revta. Fac. Ciencias Vet. U.C.V. Vol. XXVII, Nos. 1-18. pp. 11-42. 1977 -1978. 16. LEE, E. R.,TRASLEY,J., DWIVEDI,S. and LEBLOND, C. P. Divition of the mouse gastric mucosa into zymogenic and mucous region on the basis of gland features. Am. J. Anat. 164(3) : 187 -202. 1982. 17. LUNA, LEEG. Manual of histologic staining methods oftheArmed For- ce Institute of Pathologic. Lee G. Luna (Editor). Third Edition. New York. Mc Graw-Hill Book co. 258 p. 1968. 18. MENGHI, G., BONDI, A. M. and MATERAZZI, G. Correlative ultrastructural and histochamical study on submandibular gland of the hare, Lepus europpeaus. J. Anat. 139: 565-577. 1984. 19. PASHOV,V. 1981. Histología e histoquímica del duodeno del Tepez- cuintle (Cuniculus paca). Ciencias Veterinarias. Costa Rica. 3(1): 17-23. 1981. 20. REYNOLDS, J. E. and KRAUSE, W. J. A note the duodenum of the West Indian Manati (Trichechus manatus), with emphasis on the duodenal glands. Act. Anat. 114: 33-40. 1982. 21. SAKATA, T. and ENGELHARDT, W. Luminal mucin inthelarge intestine of mice, rats and guinea pigs. Cell Tissue Research. 219: 629-635. 1981. 22. SANDERSON, IVAN. Los mamíferos. Trad. F. García de Cid. Edit. Seix Barral S. A. p. 168. 23. STINSON, A. W. and CALHOUN, M. L. Sistema digestivo.In: Dellmann & Brown. 1976. Histología Veterinaria. Zaragoza, España. Edit. Acribie. pp. 223-283. 1976. 24. SUGANUMA, T., KATSUYAMA, T., TSUKAHARA, M, TATEMATSU, M., SAKAKURA, and NURATA, F. Comparative histochemical study ofali- mentary tracts with special reference to mucous neck cells of the sto- mach. Am. J. Anat. 161: 219-238. 1981. 25. TARPLEY, R., SIS R., ALBERTT., DALTON L.and GEORGE J. Observations on the anatomic of the stomach and duodenun of the bowhead whale, Baleana mysticetus. Am. J. Anat. 180: 295-322. 1987. 26. TITKEMAYER, CH. and CALHOUN, L. A comparative study of the struc- ture of the small intestine of domestic animals. Am. J. Vet. Res. 16: 152-157. 1955. 27. TROYER, HENRY. Principies and techniques of histochemystri. Little, . Brown and Company. Boston. 431 p. 1980. 28. WEIDMANN, KARL. 1979. Fauna de Venezuela. Estafeta Dorado, Panamá. Lebrería Alemana Oscar Todtmann. p. 49. 1979. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||