|
Veterinaria Tropical 16: 29-41. 1991 ESTUDIO DE PECES AUTÓCTONOS CON MIRAS A LA HIPOFIZACION S. Sette de Uzcátegui* *Universidad
de Los Andes. Facultad de Ciencias |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
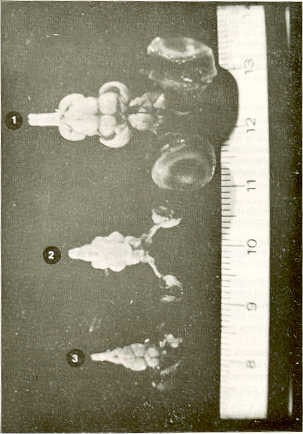
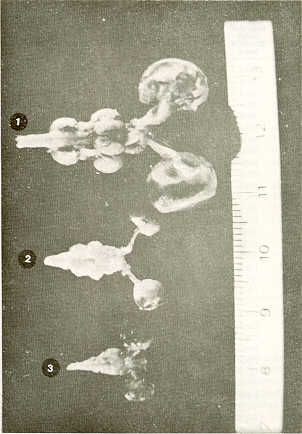
RESUMEN En la época actual, crítica para la alimentación en Venezuela, se hace necesaria una piscicultura racional que implica una reproducción controlada y el suministro de extractos hipofisiarios crudos de peces, provenientes preferiblemente de especies nativas. Sobre este particular existen pocos estudios relativos a las especies venezolanas. El objetivo del presente trabajo fue el de investigar la posibilidad de usar las hipófisis de dos especies nativas de los ríos Uribante-Gaparo. en la parte alta de los Andes occidentales de Venezuela. Se disecaron ejemplares ,de bagre Rhamdia humjlis y volador Lebiasina erythrinoides, recolectados en el río Uribante en la misma época Se tomaron varias medidas y analizaron los datos estadísticamente y posteriormente se compararon entre especies, agregando a ellas la de la carpa Cyprinus carpio. Se concluye que por las favorables características encontradas a nivel de la hipófisis, sería recomendable la explotación de Rhamdia humilis para los trabajos de reproducción inducida. INTRODUCCIÓN De acuerdo con la gran importancia que se le da mundialmente a la proteína de origen íctico y a la necesidad actual del país de encontrar y desarrollar en nuestro medio todo lo que puede satisfacer las necesidades de la población, se ha creído conveniente dirigir estudios hacia especies ícticas autóctonas que muestren algún indicio de ser rentables. Frente a la evidente necesidad de promover e incrementar la piscicultura en nuestro país, ya la importancia de la técnica de hipofización para conseguirlo, el objetivo del presente trabajo ha sido el de clarificar datos y conocimientos básicos necesarios, sobre dos especies autóctonas de la región alta de los Andes, con el fin de adaptar la técnica a nuestros medios y necesidades. Se incluyó también el estudio de una especie introducida la cual se tomó como referencia. También en los peces la reproducción es un fenómeno cíclico (11). En estas especies animales, en ambientes completamente estables, el ciclo reproductivo puede repetirse continuamente durante el período fértil de los individuos, con un ritmo determinado por el tiempo necesario para que cada ciclo se complete. Sin embargo, esta estabilidad de condiciones no se encuentra en la naturaleza, porque en ella el medio interfiere con sus continuos cambios. Los más importantes de éstos se repiten más o menos regularmente, actuando en una determinada población como marcadores de tiempo, estimulando o inhibiendo fases específicas de la gametogénesis o del proceso reproductivo en general (11 ). Los individuos que maduran sexualmente sin la acción sincronizadora de los marcadores de tiempo estarían fuera de fase frente a los cambios ambientales ya la maduración de los demás individuos de la población y no serían útiles para la especie. Cuando el pez alcanza la madurez sexual, las hormonas sexuales (esteroides) y las de la hipófisis (péptidicas) lo sensibilizan aún más a los estímulos externos (5). Importantes son la fluctuación de la luz y la de la temperatura en relación con la duración relativa del día y de la noche y con los cambios estacionales. Es evidente sin embargo, la intervención en la naturaleza de otros varios estímulos diferentes que pueden alterar notablemente el significado de "El Marcador" que se está investigando aisladamente en el laboratorio. Así, parece que en las zonas templadas, para varios Teleósteos, que son poiquiloternios, los ciclos regulares de cambio de temperatura sustituyen al fotoperíodo. Según De Vlaming (2) hay diferencias entre las varias especies respecto a los estímulos ambientales necesarios para que se complete el ciclo reproductivo normal (9). Desafortunadamente, sólo ha habido unas pocas tentativas de analizar estos requerimientos en una especie determina- da. Poco se sabe con seguridad de los marcadores de tiempo en los Teleósteos tropicales. Los estímulos captados por los órganos receptores se transforman en mensajes nerviosos que llegan al cerebro. Desde el hipotálamo los estímulos llegan a la hipófisis, a su porción nerviosa ya la glandular. La neurohipófisis de los peces es más simple que la de los vertebrados terrestres (5, 10, 11 ). El lóbulo neurohipofisiario presenta amplia interdicción en el pars intermedia (meta-adenohipófisis), alrededor del plexo capilar arterioso primario longitudinal del centro de la neurohipófisis. Aquí son liberadas las hormonas hipotalámicas que siguen hasta la adenohipófisis a través de los capilares. El mensaje de respuesta de la adenohipófisis (hormonas) será recolectada desde los capilares a unas pocas venas, llegando así a las gónadas a través de la sangre. Las gónadas, según sea el tipo de mensaje, adquirirán un determinado comportamiento a favor o en contra de la producción de hormonas sexuales o esteroides (1, 5, 8). Recientemente se está dando importancia a la glándula pineal (como componente del complejo pineal-hipotalámico-pituitario) para la regulaaión de la reproducción. La hembra de los peces requiere de un cierto tiempo para completar su maduración gonadal, pasando por cuatro fases. Durante la primera fase, la de crecimiento, se acumulan las proteínas y los lípidos necesarios en los oogonios. La duración de esta fase depende principalmente de la temperatura y de la alimentación (3, 15). En la segunda fase, o de la primera meiosis, de corta duración, hay una reorganización de los materiales, aparentemente en dependencia de la temperatura (15). Los precursores del vitelo son sintetizados en el hígado, aparentemente bajo la acción de las hormonas tiroideas (3) , mientras que la vitelogénesis depende de la presencia de la gonadotropina hipofisiaria y de los esteroides sexuales, cuyos receptores son las células tecales de la envoltura folicular. La tercera fase, o de la diapausia, encuentra a los ovarios listos y requiriendo sólo los estímulos ambientales para desencadenar la reacción de desove. Si estos estímulos no se presentan, los oogonios pierden la capacidad de madurar ulteriormente y se reabsorben atreticamente. También la duración de esta fase parece depender de la temperatura y de la alimentación. La cuarta fase, o de segunda meiosis, es la fase de maduración final. Está dirigida única y exclusivamente, por las hormonas sexuales, por lo que no puede ser detenida ni retardada con otros medios (15), siendo la 17-Hidroxi-20 dihidroprogesterona (17 a-20ß Pg) el mediador más probable (7, 13). Al terminar esta fase se produce la reabsorción del agua, la ruptura folicular y la separación de los oocitos de la pared del ovario y su caída en la cavidad del mismo, de donde son impulsados a los conductos que lo llevarán hacia el exterior (3, 15). Según varios investigadores (3, 7),la ovulación parece ser independiente del control hipofisiario, siendo los mediadores las prostaglandinas y las catecolaminas. Las hembras de varias especies, en cautiverio, llegan solamente a la tercera fase de maduración sexual por faltar los estímulos naturales ambientales de importancia, como por ejemplo, los estimulantes olfatorios, traídos por el agua corriente (11). En muchos Teleósteos la ovulación no ocurre espontáneamente, aun en presencia del macho (14). Sin embargo, con la técnica de hipofización se puede favorecer la maduración sexual completa y la expulsión de los productos genitales, consiguiendo la reproducción. Por lo general, la maduración gonadal de machos en cautiverio no se ve interrumpida. En él se aplica la técnica de hipofización para asegurar la coordinación de la liberación de los gametos y la mejor calidad del esperma. Con la técnica de hipofización se estimula la producción de las hormonas sexuales bajo el estímulo de las hormonas hipofisiarias. La hipofización consiste en inyectar intramuscularmente, por lo general, o intraperitonialmente, extractos crudos de hipófisis de pez. La actividad de estos extractos se relaciona con la distancia filogenética entre el donante y el receptor; el sexo, la edad, la madurez sexual del donante, el método de extracción de las hipófisis y la técnica de su preservación. La técnica de hipofización implica un cierto trabajo y tiempo, pero si se tratan debidamente los extractos (deshidratación y envasado estéril) se pueden almacenar indefinidamente para posterior utilización. No se puede llegar a standarizar la dosificación sin conocer previamente la actividad gonadotrópica de los extractos. La distancia filogenética entre donante y receptor es muy importante puesto que los extractos heteroplásticos requieren dosis mayores. MATERIALES Y MÉTODOS En relación con las técnicas de hipofización se estudiaron y compararon dos especies autóctonas de los Altos Andes Venezolanos: bagre capitanejo (Rhamdia humilis) y volador (Lebiasina erythrinoides) por ser de explotación potencial industrial, deportiva y de promoción rural en la zona de los embalses del Complejo Hidroeléctrico en el occidente venezolano, sobre los ríos Uribante-Corados y Caparo-Camburito. También se consideró una especie exótica introducida y bien estudiada en Venezuela: carpa (Cyprinus carpio). En ejemplares de las dos especies autóctonas recolectados en la misma zona y en la misma época, se enfocó un estudio anatómico con mira a la técnica de hipofización (12). Se realizaron disecciones directamente en especímenes preservados en formol al 10% y sin utilizar maceración. Previa disección del cráneo y del sistema nervioso central con sus anexos olfatorios y ópticos, se disecó la región hipofisiaria de los especímenes desde el interior del cráneo, cuidando de obtener la hipófisis completa y en buenas condiciones. Se realizaron las mediciones de peso y de dimensiones sobre los tres planos de cada hipófisis obtenida; los datos fueron procesados estadísticamente. RESULTADOS La hipófisis se encuentra en la base del cerebro, debajo del hipotálamo e inmediatamente detrás del quiasma óptico, encajando en un nicho óseo del piso craneal, la silla turca. El tejido hipofisiario está rodeado y protegido por una prolongación de la membrana meníngea primaría que también sella el nicho superiormente, rodeando el paso del pedúnculo hipofisiario. La morfología de la hipófisis y de los tejidos que la rodean presenta diferencias de importancia entre las tres especies estudiadas (Figuras 1 y 2). En el volador y en la carpa el plano entre la hipófisis y el techo nasal prácticamente no existe, estando constituida la separación por la meníngea primaria, y en el caso de la carpa, por un cierto espesor de tejido conjuntivo. También se encuentran diferencias en la membrana meníngea primaria; en el volador se presenta como una capa fibrosa dura que dificulta bastante la extracción de la glándula; en el bagre y en la carpa es notablemente más suave y frágil, lo que facilita la disección. En las tres especies, el pedúnculo de la hipófisis se inclina hacia atrás. Una vez extraída la hipófisis de su nicho óseo se observa que éste es poco evidente y prácticamente plano en el bagre. En el volador la concavidad es ligera pero evidente, y en la carpa el nicho es más profundo y evidente. En el bagre la hipófisis, vista desde arriba, es prácticamente redonda, pre- sentando una forma de pera en la vista lateral y disminuyendo progresivamente su grosor hacia atrás. En el volador se observa una estructura piriforme en una vista superior y elíptica regular desde una vista lateral. En la carpa (Cyprinus carpio), la forma es similar a la encontrada en el bagre. En el bagre, las medidas relativas de la hipófisis son mayores que en las otras dos especies (Cuadro 1) ; le sigue la carpa, en la cual el ancho relativo de la glándula es algo mayor que en el volador, siendo casi iguales las otras medidas relativas (Cuadros 2 y 3).
Por lo que se refiere a los valores absolutos de volumen y peso, éstos son mayores en la carpa, siguiendo el bagre y por último el volador. Sin embargo, el peso de la hipófisis relativo a las medidas del ejemplar, largo standard (LS), son mayores en el bagre. En el Límite inferior común de medidas (LS) el peso relativo es menor y el mismo para el volador y la carpa. Sin embargo, en el límite superior de medidas (LS) el peso relativo de la hipófisis de la carpa supera a el del volador, aunque quedando inferior a el del bagre. Para el volumen, se observa una relación paralela. La asociación entre el volumen y el peso de la hipófisis y (LS) es mayor en la carpa y algo menor y equivalente entre el bagre y el volador. La prueba "t" de Student demostró que dichas asociaciones son significativas (probabilidad de P=0,001).
DISCUSIÓN Considerando las especies estudiadas, se puede señalar que la silla turca presenta una morfología diferente en las tres especies, siendo más simple en el bagre, en el cual hay además una separación ósea de importancia entre la hipófisis y las fosas nasales. Por ello, la extracción de la hipófisis es más rápida y fácil en el bagre. La mayor resistencia de la membrana meníngea primaria sobre la hipófisis en el volador, asociada al menor tamaño promedio de la especie, dificulta en ella, más que en las otras dos especies, la disección y extracción de la hipófisis. Las medidas relativas de la hipófisis, en relación al (LS) es diferente en cada una de las tres especies estudiadas, siendo mayores en el bagre. Evidentemente, si se utilizara la técnica de hipofización, al utilizar la hipófisis de bagre, por el peso absoluto y relativo de ésta, se requerirá de un menor número de glándulas para preparar los extractos. CONCLUSIONES Generalmente para la hipófisis se han utilizado extractos hipofisiarios importados de carpa. Como esto resulta actualmente muy costoso y deja la inseguridad en relación con su calidad (no pudiendo excluirse la posibilidad de que sean causa de problemas ictiopatológicos) , el uso de extractos de hipófisis de especies autóctonas sería muy importante, recomendable y conveniente. En nuestro caso, entre las dos especies autóctonas estudiadas, se deberían utilizar los del bagre, ya que el peso de su hipófisis, sea en sentido absoluto como relativo es mayor en comparación con el del volador. Por otra parte, como se mencionó, la extracción de la hipófisis en el bagre es más fácil y más rápida que en el volador. Lo que queda por verse en futuras investigaciones es cómo actúan los extractos a diferentes dosis, y que especies receptoras se benefician más para la inducción. Sería recomendable utilizar los extractos en las mismas especies, evitando los problemas por la distancia filogenética; como también investigar si los extractos actúan mejor solos o combinados (alternados) con gonadotropina coriónica humana (GCH); ya que ha sido comprobado que en ciertas especies como la carpa de la China y de la India, este método resulta efectivo y económico (extractos hipofisiarios heteroplásticos u homoplásticos). SUMMARY With the decreasing supply of essential food, specifically in Venezuela, more attention should be paid to fish as a protein source, to rational fish hatcheries, to species which are presently little used for human consumption, and to hipophyzation using native species. On this respect a little information exists on Venezuelan fish species. The aim of this work is to use, in acquaculture practice, the hypophysis ot two native species of the Uribante-Caparo rivers, from the south western Andes of Venezuela. Rhamdia humilis (catfish) and Lebiasina erythrinoides (flying fish), colleted Uribante basin, were dissected and measured analyzed. and compared the two species and carpa (Cyprinus carpio) The results suggest the convenience to utilize R. humilis hypophisis in induced spawing. BIBLIOGRAFÍA
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||