|
Veterinaria Tropical 16: 69-110. 1991 CARACTERÍSTICAS ANATÓMICAS DE TRES ESPECIES DE PECES DE LA CUENCA ALTA DEL RÍO URIBANTE EN RELACIÓN CON SU FISIOLOGÍA REPRODUCTIVA S. Sette de Uzcátegui* *Universidad de Los
Andes |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
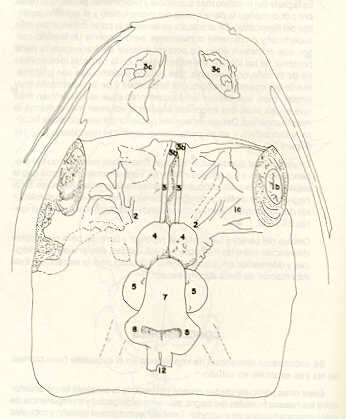
RESUMEN En la época actual se necesita de una piscicultura racional para la alimentación de la humanidad, sin embargo los estudios de las especies venezolanas son escasos La piscicultura racional implica la reproducción controlada y eventualmente la fecundación e incubación artificial, posiblemente con una fecundación mejorada por medio de los extractos hipofisiarios. El objetivo de este trabajo fue conocer la explotación de las bases anatómicas y fisiológicas de dos especies nativas de potencial, originarias de la cuenca alfa del río Uribante en el occidente montañoso venezolano Rhamdia humilis (bagre) y Lebiasina erythrinoides (volador) Adicionalmente se compararon las dos especies con una exófica Cyprinus carpio (carpa). En el laboratorio del Grupo del Ecología Animal, de la Universidad de Los Andes, Mérida- Venezuela, se estudió la anatomía fisiológica general, profundizando el estudio y la técnica de disección para la mortologia craneal, relacionada con la hipófisis Se encontraron importantes diferencias mortológicas entre las tres especies, en el sistema nervioso central y en el cráneo. En el bagre la silla turca es más simple y más osificada, la hipófisis es más grande, la extracción de la carpa pineal es más fácil y rápida hay una mayor producción de huevos y esta especie es aceptada como alimento en la cuenca alta de los Andes Por lo tanto, la piscicultura de esta especie, por siembra y extractos hipofisiarlos. tendría que ser más fácil para el uso comercial y deportivo. INTRODUCCIÓN El conocimiento anatomo-fisiológico de las especies de nuestra ictiofauna es una de las bases indispensables para el planeamiento racional de los cultivos eficientes de peces, por lo cual está adquiriendo cada vez mayor importancia para satisfacer la necesidad de alimentación de la humanidad. La mayoría de los estudios realizados hasta el momento en el país se refieren a las zonas bajas: los llanos, la cuenca del Lago de Maracaibo y la región oriental (2, 3, 4, 5, 6, 7, 8, 9, 11, 12, 13, 14, 20 y 21 ). Muy poco se ha estudiado en los Andes, salvo a lo que se refiere a la trucha (Salmo gairdnerii, Rrchardson) por parte del Fondo Nacional, de Investigaciones Agropecuarias (21) y la Corporación de los Andes (CORPOANDES). Paralelamente a este estudio Pérez*, en 1986, ensayó el desarrollo embrionario y juvenil del bagre (Rhamdia humilis), y ya en 1978 Machado-Allison (10) había estudiado el desarrollo del volador (Lebiasina erythrinoides). El objetivo de este trabajo fue conocer las bases anatómicas y fisiológicas de dos especies nativas, el bagre (Rhamdia humilis) y el volador (Lebiasina erythrinoides). MATERIALES Y MÉTODOS Las especies estudiadas son: bagre (Rhamdia humilis) y volador (Lebiasina erythrinoides) pertenecientes a la clase teleosteii. Los ejemplares de R. humilis y L. erythrinoides fueron recolectados en la cuenca alta del río Uribante, estado Mérida, en la misma zona y en la misma época. Los resultados de las observaciones se confrontaron con los relativos a una tercera especie de la misma clase: carpa (Cyprinus carpio), no autóctona y mucho más estudiada. Se enfocó el estudio anatómico esencialmente con miras a profundizar en relación con su fisiología reproductiva ya la técnica de hipofización. La observación anatómica y la disección de los especímenes se hicieron más detallados en relación con la morfología sexual, al cráneo ya la región hipofisiaria, en orden creciente. Todas las disecciones se hicieron directamente en especímenes capturados en la región indicada, preservados en formol al 10% y sin utilizar ninguna técnica de maceración previa. Para el estudio del sistema nervioso central de las especies consideradas, y tomando como prototipo para la técnica empleada al bagre, se procedió a marcar la línea de corte en la parte anterior reuniendo los bordes anteriores de las órbitas; en la parte posterior, reuniendo la inserción de los arcos branquiales de cada lado, y lateralmente, a nivel de cada lado, desde el borde inferior de la órbita, trazando una línea paralela a la línea lateral (Figura 1 ). Se realizó el corte perpendicularmente a las superficies externas correspondientes, eliminando la piel y los planos muscular y óseo. Al retirar este último se descubre el cerebro (Figura 1) y los nervios olfatorios o los pedúnculos olfatorios (No.3); quedan expuestos también posteriormente las partes superiores de las branquias. Eliminando tejidos blancos y duros hasta el nivel de la base del cerebro, se descubre el bulbo (No.1 b) y la mitad superior de los globos oculares; se eliminaron todos aquellos tejidos que impiden la visibilidad de los nervios ópticos (No.2), hasta que quedaron expuestos. Al finalizar estos pasos se pudo observar claramente la forma del cerebro y de sus componentes (Figuras 1 al 5). Los datos biométricos fueron procesados estadísticamente. RESULTADOS Y DISCUSIÓN Resumiendo los datos de mayor importancia obtenidos se tiene: - Las escamas del bagre (R. humilis) son de tipo ciclideo, son más finas, suaves y pequeñas, lo que es conveniente para el eventual comercio y consumo. Esta especie presenta también una mayor propor¿;ión de carne comestible debida a la forma del cuerpo, la cual presenta solamente un achatamiento dorsoventral moderado, mayor en la porción anterior, en contraposición al volador (L. erythrinoides) que presenta achatamiento lateral. - El tamaño del estómago sugiere que ambas especies son voraces, aunque el consumo energético relativo tiene que ser mayor en el volador debido a su gran movilidad. - De las tres especies, solamente el bagre cambia su coloración según el medio que lo rodea y los estímulos bajo condiciones de laboratorio.
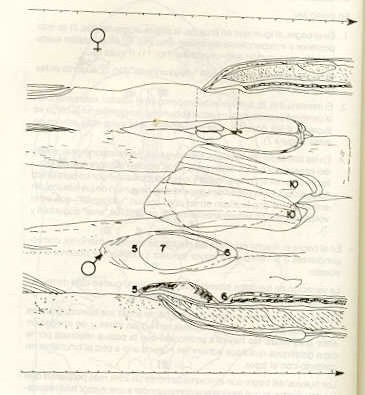
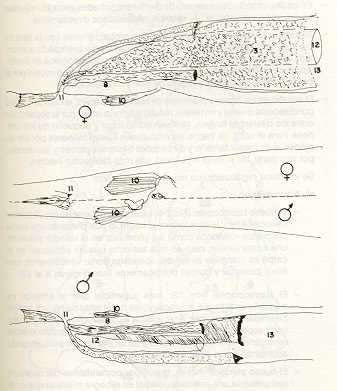
-Las tres especies presentan dimorfismo sexual. En el bagre y en la carpa , (C. carpio) la diferencia es evidente tanto en la papila urogenital como en la forma o implantación de las aletas; en el volador es menor la diferencia en relación con la papila urogenital. -En particular: 1. En el bagre, al igual que en la carpa, la papila urogenital (No.7) es más posterior e independiente del orificio anal (No.6); en el volador existe un solo poro común ano-urogenital (No.11) (Figura 6). 2. El bagre presenta una pequeña "vejiga urinaria" (No.2), ausente en las otras dos especies. 3. El intestino (No.8) , tanto en el bagre como en el volador, está situado en la parte más ventral de la cavidad abdominal, en cambio, en la carpa es superior a las gónadas sobreponiéndose a la parte más inferior de estas (Figura 7). 4. En las tres especies, los túbulos encargados del transporte de la orina y del esperma u óvulos están separados, pero en el bagre ambos túbulos se unen en la parte terminal y los productos salen por un poro único "urogenital". En las otras dos especies no hay unión de los túbulos, sin embargo, los dos finalizan en un poro común "urogenital", que en el volador, como se señaló anteriormente, es también "anal" (Figuras 6 y 7). -En el bagre el macho presenta las medias de variables menores, correspondientes a la hembra de mayor tamaño. Lo mismo se observa en el volador. La variación de los datos biométricos hace pensar que se trata de muestras representativas de las relativas poblaciones. En el área de captura se observó que el bagre anida sus huevos, grandes y rodeados por una masa gelatinosa, en aguas libres y de anegación entre gramíneas. No hay otra protección que la pasiva realizada por la capa gelatinosa que hace adherir los huevos uno a otro al hincharse en , contacto con el agua. Los huevos del bagre son aproximadamente un 25% más pequeños que los del volador, lo que tendría que corresponder a una menor tasa respiratoria y necesidad de mayores niveles de energía (17). -El bagre presenta un mayor número absoluto y relativo de huevos, lo que está de acuerdo con el menor tamaño de los mismos. Relacionando el número de huevos con la longitud standard (LS), el número de huevos producidos por el bagre resulta ser de 2,5 a 3 veces mayor que los del volador. El tamaño aproximado de los huevos del bagre es de 0,8 mm, mientras que los del volador son de 1,8 mm (2,25 veces más grandes).
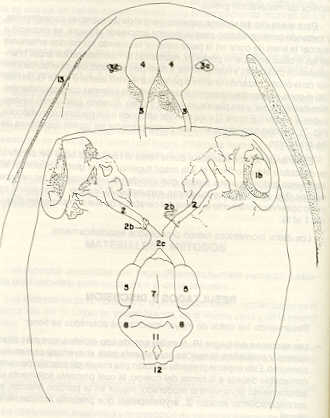
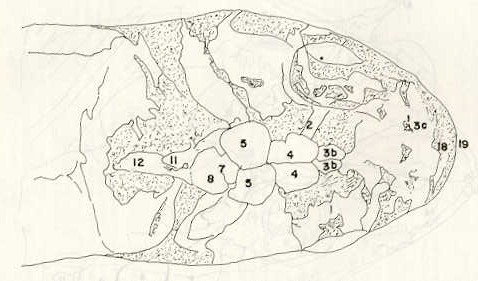
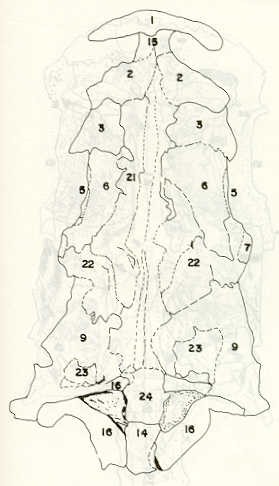
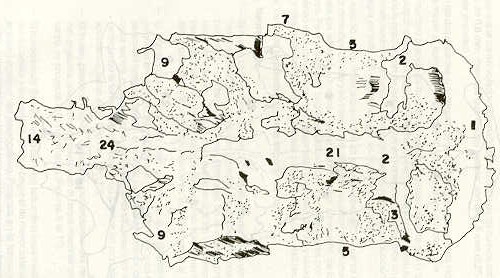
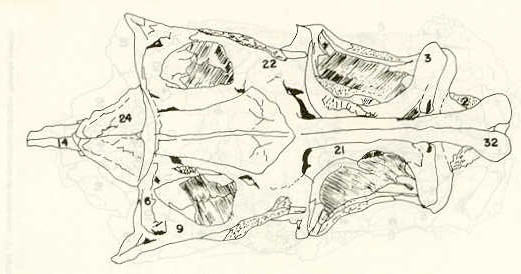
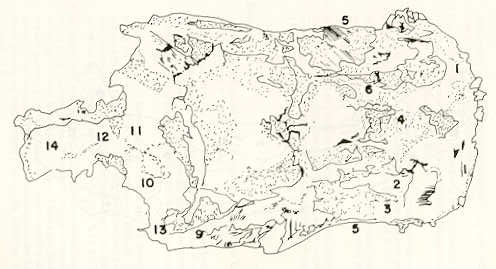
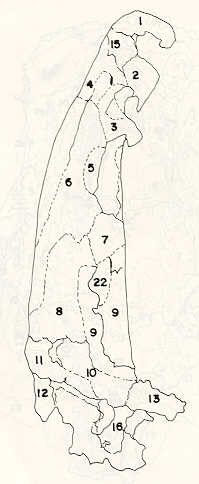
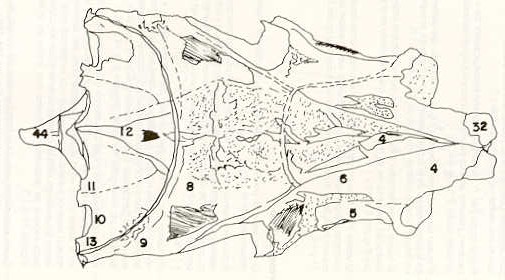
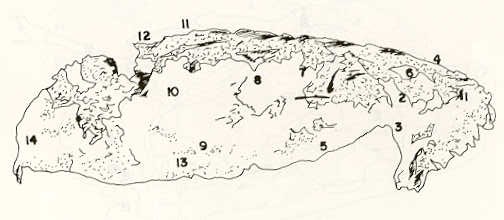
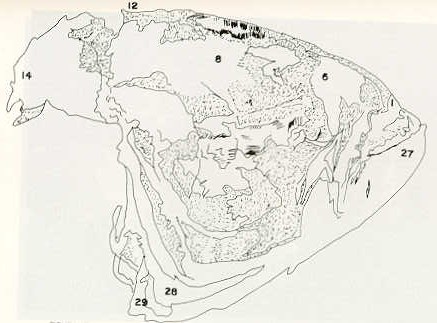
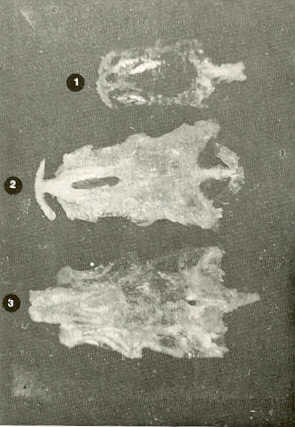
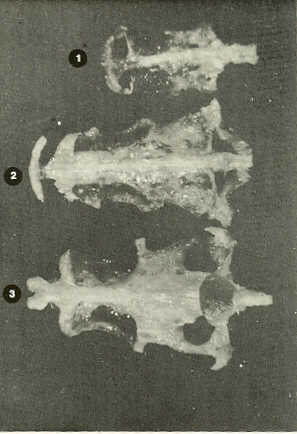
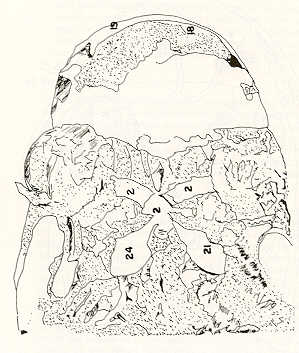
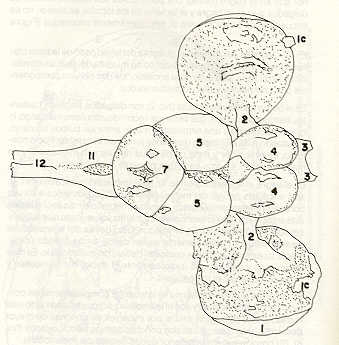
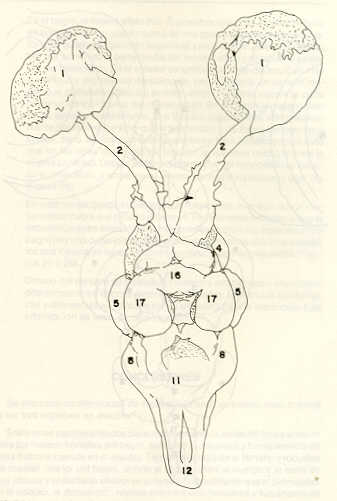
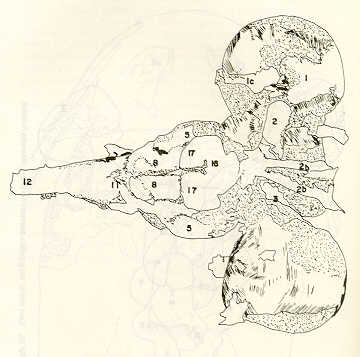
- El número crecido de huevos se relaciona probablemente con la falta de cuidado activo, pudiéndose esperar que en un medio controlado aumentaría la producción de larvas y alevines frente a lo que acontece en condiciones naturales, resultando entonces mucho mayor la producción por el bagre en vista del mayor número de huevos producidos. - El estudio de los huesos craneales de las tres especies, el cual se realizó por disección directa y en fresco, resultó difícil visualizar por la separación entre los diferentes huesos, también por lo frágil y pequeño de los cráneos. Para el caso del bagre, las dificultades fueron menores por ser su cráneo el de mayor tamaño y de paredes bastantes planas y resistentes; por otra parte, las suturas intraóseas son más fácilmente visibles. De los datos registrados, los más importantes son: a) Neurocráneo - El hueso basioccipital (No.24), localizado delante de las vértebras cervicales (No.1) es mayor en el bagre, el cual presenta una forma prismática-cilíndrica como las vértebras; en el volador presenta una forma similar, pero es más estrecho que las vértebras; en la carpa es triangular piramidal, aplastado dorso ventralmente, con ápice posterior y bordes prácticamente lisos (Figuras 8 al 13). - El supraoccipital (No.12), más superficial que el anterior, es pequeño en el bagre, proyectándose hacia atrás como una espina piramidal ovalada libre; en el volador es de tamaño menor, con una corta espina posterior; en la carpa presenta un desarrollo notable recordando la forma de fa cabeza de un ave con un pico alargado apuntando hacia atrás (Figuras 8 al 13). - El hueso pterótico (No.9), hueso par posterolateral del neurocráneo presenta una extensión relativa en el bagre mientras que en la carpa se proyecta más hacia atrás (Figuras 8 al 15). - El hueso frontal (No.6), hueso par a los lados de la línea media anterior al parietal y al pterótico, es en el bagre el de mayor extensión en la pared superior del neurocráneo. Participa en la formación del techo de la órbita. En su mitad anterior, en correspondencia con la línea media del neurocráneo, se ha encontrado falta de calcificación, presentándose una ranura bastante ancha que permite visualizar el interior del cráneo (Figuras 8 al 17).
- El post frontal (No.7) , hueso par pequeño lateral posterior del neurocráneo, presenta en la carpa una corta espina dirigida hacia atrás de base ancha (Figuras 8 al 15). - El hueso lacrimal o paratmoides (No.3), localizado lateralmente en la parte anterior de la órbita y anterior a los frontales, presenta un mayor tamaño relativo en la carpa (Figuras 8 al 17). - El hueso paraesfenoides (No.21), en la porción central anterior ventral del neurocráneo es relativamente más extendido en el bagre (Figuras 8 al 10 y 14). - El vómer (No.15) se proyecta en el bagre entre el premaxilar anteriormente y el paraesfenoides posteriormente, siendo visible, en el cráneo óseo, tanto dorsal, ventral como lateralmente (Figuras 8, 9, 11 y 14). b) Branquiocráneo - El arco visceral, posterior a los maxilares, se observa bastante desarrollado en el bagre. - El maxilar (superior) (No.12), hueso par, es voluminoso en el bagre, localizándose delante del lacrimal, al mesetmoides (No.4) y al frontal, posteriormente a la porción visible del vómer y lateralmente a la porción anterior del paraesfenoides en la cara ventral (Figuras 8, 9 y 11 al 14). - Premaxilar (No.1), hueso par, ubicado delante del maxilar y del vómer (No.15). En el bagre, con el contralateral, adquiere forma de boomerang, con sus porciones laterales prácticamente suspendidas delante del cráneo óseo. Únicamente su parte central se une al cráneo por intermedio del vómer (Figuras 8, 9, 14 y 15). - El dentario mandibular (No.27) , hueso par que cubre el borde de la mayor parte del maxilar inferior, presenta un gran desarrollo tanto en el bagre como en el volador, proyectándose por delante del maxilar superior y siendo el hueso que más se destaca en la vida lateral. Sin embargo, para las mismas características del cráneo de cada especie es relativamente mucho más ancho en el bagre (Figuras 15 al 17). - Desde el punto de vista de la forma general del cráneo, las tres especies se diferencian notablemente (Figuras 18a, 18b ).
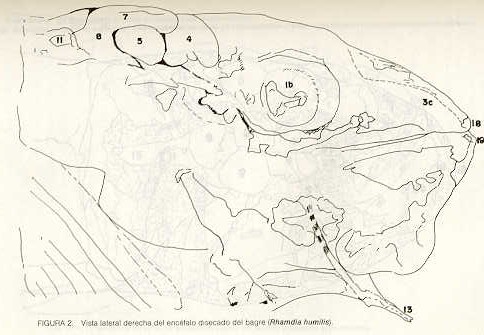
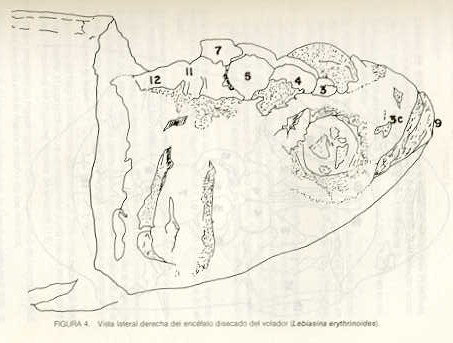
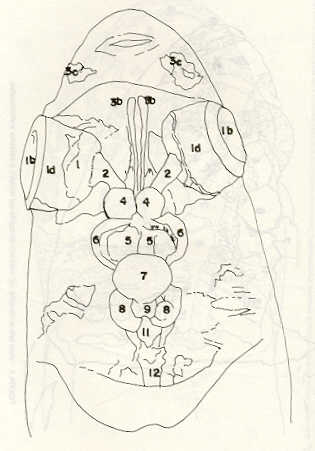
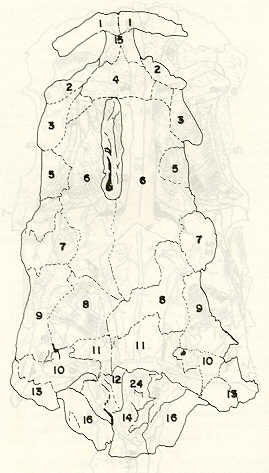
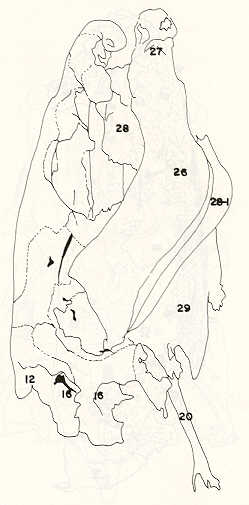
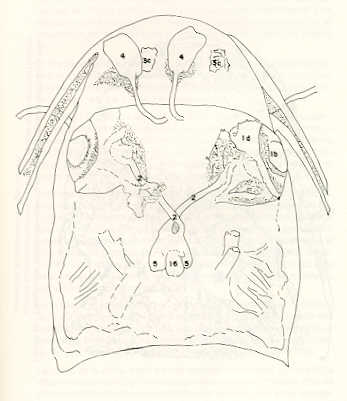
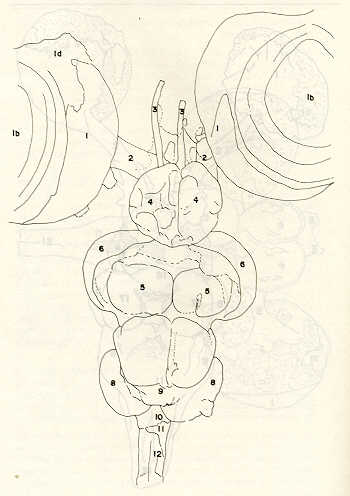
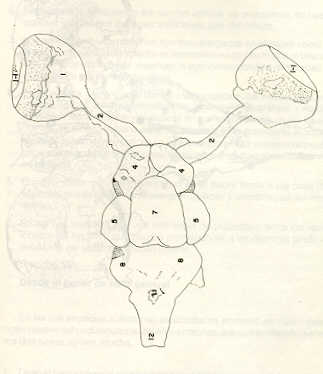
Igualmente es notable la diferencia de forma y proporción del maxilar inferior y del dentario entre el bagre y el volador, así como la marcada ranura entre las mitades anteriores de los frontales en el bagre. También se diferencia en la carpa la forma de supraoccipital. Además, en esta especie el límite posterior superior del cráneo es relativamente más anterior y cubierta por tejido muscular y piel con escamas. Debido a su resistencia el cráneo del bagre se puede manipular más fácilmente, con menor peligro de rupturas que pue den perjudicar el posterior estudio anatómico y topográfico del encéfalo. c) Sistema nervioso Debajo del telencéfalo se localiza el quiasma óptico (No.20) , el cual en la gran mayoría de los teleósteos están formados por el cruce de los nervios ópticos, sin ninguna asociación de sus fibras (15, 16) .En con secuencia, los dos nervios ópticos pueden disecarse por separado, aun en correspondencia del quiasma. Este es el caso de las tres espe cies en estudio, desplazando hacia adelante los lóbulos cerebrales anteriores con los pedúnculos olfatorios, se descubre el quiasma Ópti co (Figuras 19 y 20) . Según estudios anteriores de diferentes especies de teleósteos (15), en la misma especie se encuentra con frecuencia un porcentaje variado entre las dos posiciones relativas posible de los nervios ópticos. En consecuencia, se examinó al respecto una porción de las muestras estudiadas. Para el bagre se obtuvo los siguientes porcentajes: posición dorsal del nervio óptico derecho 76% y posición ventral del nervio óptico derecho 24%, para el volador, posición dorsal 66,67% y para la carpa 45,45%. Así, en ejemplares estudiados de las especies autóctonas, predomina la porción dorsal del nervio óptico derecho, lo que está de acuerdo con la observación de Parker (15) , para 10 especies de teleósteos. Sin embargo, la apreciación del porcentaje obtenido tendrá que someterse a un posterior estudio específico con un mayor número de ejemplares eventualmente diferenciado los resultados para los dos sexos. Los lóbulos olfatorios (3b) del bagre se alejan bastante hacia adelante, llegando a nivel de la pared anterior de la órbita; son pequeños y se conectan al telencéfalo (4) por largos y delgados pedúnculos olfatorios (3) (Figura 2). En el votador los lóbulos olfatorios están prácticamente en contacto con el telencéfalo, localizándose a nivel de la porción media de la órbita. Su tamaño relativo es algo mayor que en el bagre (Figuras 4 y 5). En ambas especies los dos lóbulos están en contacto uno con el otro.
En la carpa, al igual que en el bagre, los lóbulos olfatorios se encuentran distanciados del telencéfalo, a un nivel relativo un poco más anterior que en el bagre mismo. Los pedúnculos son relativamente más delgados que en el bagre y el tamaño de los lóbulos es menor; no se encuentran en contacto entre sí, sino ligeramente separados (Figura 19). En el volador es difícil liberar el quiasm a del tejido óseo de la base craneal que lo engloba. El quiasma óptico no muestra diferencias en relación con el sexo, y es externo al encéfalo; los dos nervios componentes son compactos y bien individualizados. En el bagre los nervios ópticos (No.2) son delgados, frágiles y bastantes alargados, es decir, que tienen un recorrido intra óseo más largo, lo que corresponde a una separación mayor entre los bulbos oculares, resultando más dificultoso disecarlos junto con el cerebro. Por lo contrario, en el volador y en la carpa estos nervios son más cortos y de mayor grosor. La carpa es la que presenta mayor grosor. En el bagre, el mesencéfalo (No.5) , justo detrás del telencéfalo (No.4) , está encajado debajo del metencéfalo (No.7), cuyos lados a su vez bordea. Presenta forma elíptica, siendo su superficie más ancha la que está en contacto con el telencéfalo. Es menos voluminoso que el telencéfalo o el metencéfalo. En el volador, justo detrás del telencéfalo el mesencéfalo es completamente visible desde arriba, siendo Únicamente su extremo posterior encajado debajo del metencéfalo. Es más voluminoso y más grande (prácticamente el doble) del telencéfalo y un tercio más que el meténcéfalo. En la carpa, el mesencéfalo no se encuentra completamente en contacto con el metencéfalo y con el telencéfalo, porque en cada lado está rodeado, de abajo hacia arriba, por especies de conchas de mayor desarrollo anterolateral. Las dos conchas son los tractos ópticos (No. 6). Su porción extrema posterior queda debajo del metencéfalo. La carpa es la única, entre las tres especies en estudio, que presenta esta formación de fibras nerviosas tan evidente y diferenciable. En esta especie, el tamaño del mesencéfalo en sí es menor que el del telencéfalo o del metencéfalo, pero, en conjunto con las conchas laterales, viene a ser la parte del encéfalo más grande y voluminosa. (Figuras 21 al 23). Como regla general, en los teleósteos los lóbulos ópticos y los bulbos oculares están dispuestos simétricamente, independientemente del cual sea el curso del nervio óptico (15).
En el bagre, el metencéfalo (No.7) presenta una gran extensión, tanto en sentido anteroposterior como en una porción posterior transversal. Es la parte del encéfalo más superficial y prominente, puesto que recubre por completo la parte media del mesencéfalo y el extremo posterior del telencéfalo. En el volador es también la parte del encéfalo más superficial y prominente dorsalmente, pero presenta un tamaño menor que el mesencéfalo. Su porción anterior central separa la parte posterior de los lóbulos del mesencéfalo. En la carpa, el metencéfalo es de tamaño considerable. Sus lóbulos superficiales son prácticamente del mismo tamaño que el telencéfalo y de forma también muy similar, pero sus lóbulos lateroposteriores (que se diferencian más que en las otras dos especies) añaden al volumen más o menos la misma cantidad. Debajo de la mitad posterior del telencéfalo se localiza el diencéfalo, y en su parte inferior central el hipotálamo (No.16) (Figura 24) En vista dorsal queda cubierto por el telencéfalo, mientras que el mesencéfalo cubre sus lóbulos inferiores. Ventralmente se aprecia que la separación entre estos lóbulos inferiores varía entre las especies. En el bagre hay una distancia notable entre ellos, mientras que en el volador los dos lóbulos se tocan en correspondencia con la línea media (Figuras 25 y 26). Debajo del cerebro se localiza la hipófisis y también para ella existen diferencias entre las tres especies estudiadas, diferencias morfológicas y diferencias en relación con la dificultad para la extracción. Esta información es tema de otro trabajo. CONCLUSIONES Se encontraron diferencias de importancia en el esqueleto óseo craneal de las tres especies en estudio. Entre otras particularidades parece interesante la evidente fisura anterior entre los huesos frontales del bagre, así como la delgadez y transparencia de estos mismos huesos en el volador. También es notable el tamaño y robustez del maxilar inferior del bagre, donde el ángulo entre el cuerpo y la rama es muy obtusa y el dentario inferior se proyecta más adelante que el premaxilar. En el volador, la disposición relativa entre los dos maxilares y sus anexos es parecida, pero el ángulo mandibular es mucho más agudo, de acuerdo con las mayores dimensiones relativas verticales frente a las transversales. En esta especie es también notable la agudeza de los dientes.
Por lo que se refiere al sistema nervioso central: 1. El nervio óptico del bagre es notablemente más largo que el de las otras dos especies en relación con la forma de la cabeza de esta especie que es nada en sentido vertical, aumentando la dimensión transversal y por la distancia entre los bulbos oculares. Por otra parte, en esta especie los nervios ópticos son más delgados y frágiles. 2. Respecto a la posición de los nervios ópticos en el quiasma, se puede concluir que las dos especies autóctonas son dimórficas. 3. Los nervios y lóbulos olfatorios son muy alargados en el bagre como en la carpa, mientras en el volador se mantienen en contacto con el telencéfalo. Sería de gran interés investigar si esto corresponde a diferencias en el desarrollo del sentido del olfato. 4. En la carpa, a diferencia de las otras dos especies, los trechos ópticos y nervios ópticos son muy prominentes, fácilmente separables y visibles, , continuando los nervios ópticos en correspondencia de los lóbulos ópticos, se dirigen hacia la parte del encéfalo. Esto acompaña al notable espesor de los nervios ópticos en esta especie, y sería interesante investigar si corresponde a alguna característica o especial agudeza de la visión en la carpa carpa. 5. El mayor desarrollo del metencéfalo en el bagre frente a las otras dos especies, puede relacionarse con gran agilidad y coordinación del movimiento. 6. En las tres especies hay una diferencia de proporción entre los varios lóbulos del encéfalo que podrán corresponder a un diferente grado de lución del sistema nervioso central. Desde el punto de vista general En las dos especies autóctonas estudiadas se encontró un mayor desarrollo relativo del pedúnculo caudal en los machos, aunque la diferencia entre sexos no sea mucha. 1. Tanto el bagre como el volador presentan dimorfismo sexual, lo que facilita el reconocimiento de los reproductores, pero este reconocimiento es más rápido en el bagre. 2. La talla de la especie estudiada no es muy grande. Sin embargo, el tamaño alcanzado por el bagre es superior al del volador y comparable el de otras especies actualmente en el comercio. 3. El bagre es voraz, pero parece tener un menor consumo energético. 4. El bagre produce un mayor número de huevos, por lo tanto, con un adecuado cuidado de incubación se podría obtener un mayor número de alevines por cada desove. La explotación del bagre sería recomendable porque puede ser sembrado y adquirir un valor comercial y deportivo de importancia; se trata de una especie nativa, conocida y aceptada, a nivel de consumo, por los habitantes de las cuencas altas de los andes. SUMMARY At the present critical time, a national fish culture is necessay for the mankind nourishmenl Until now, the studies of the Venezuelan species are only about a few. The national fish culture implicates the controled reproduction and, eventualy, the artificial fecundation and incubation, possiblywith a improved fecundation by means ofthe hypophysis extracts. The objetive of this work was knowing the anatomic and physiologic basis of two natlve species of potential exploitation: Rhamdia humilis Capitanejo (catfish) and Lebiasina erythrinoides (Flying fish). We compared the two species with a foreign one: Cyprinus carpio (carp).ln the laboratoryofthe Grupo de Ecología Animal -Universidad de los Andes, Mérida -Venezuela, was studied the general anatomy physiology, deepened the study and the dissection tecnic for the cranial morfology related with the hipophysis. The study gots important morfologycal disimilarities between the three species in the central nervous system and in the craneal skeleton. In the catfish the "Sella Turcica" is more simple and more ossified, the hypophysis is bigger; the extraction ofthe pineal body is easier and faster; there is a production of mo(e eggs, and this specie is accepted as food in the high basins of the Andes. For that, the fish culture of this specie, by deeding and hypophysis extract, would be easier for the commer cial and sport use. BIBLIOGRAFÍA 1. BERMUDEZ, D., N. PRADA y C. KOSSOWSKI. Ensayo sobre la reproducción de cachama (Colossoma macropomus) (Cuvier) 1818, en cautiverio. Barquisimeto, Ven. Universidad Centro. Occidental, Escuela de Agronomía. Dirección Extensión Universitaria. 1979. 23 p. 2. CORPORACION DE LOS ANDES. Programa truchícola. Evaluación preliminar del recurso agua del estado Mérida con fines de explotación truchícola. Mérida, Ven. 1988. 290 p. 3. ESPINOSA, U. y G. GIMENEZ. Estudio de la biología y pesca del bocachico (Prochilodus reticulatus) (Valenciennes) en el Lago de Maracai bo. Caracas. Ministerio de Agricultura y Cría. Informe Técnico No. 63. 1974.30 p. 4. GARCIA, P., L. QUIÑONEZ y G. GONZALEZa. Biología reproductiva de wHypostumus watwata (Osteicithye loricariidae), armadillo pintado del Lago de Maracaibo, Venezuela. Carrasquero, Ven. Centro de Aprendizaje "Don Bosco". Boletín Técnico No.3. 1984.20 p. 5. KOSSOWSKI, C. Ensayo de reproducción inducida en palometa carachica (Mylossoma duriventris) (Cuvier 1818) (Pisces, Cypriniformes) con el uso de gonadotropina coriónica humana. Acta Cient. Venezolana 31: 444-448. 1980. 6. KOSSOWSKI, C. y F. MADRID. Ensayo de reproducción inducida del bagre rayado cabezón (Pseudoplatystoma fasciatum) (Linnaeus). Acta Cient. Venezolana 36: 284-285. 1986. 7. KOSSOWSKI, C., J. VALDES y N. PRADA. Ensayo sobre hibridización artificial entre cachama (hembra) (Colossoma macropomus) (Cuvier) y palometa carachita (macho) (Mylossoma durlventris (Cuviér) 1818. Barquisimeto, Ven. Escuela de Agronomía. Universidad Centro occidental Lisandro Alvarado. 1980. 30 p. 8. LILYESTROM, C. Aspectos de la biología del coporo (Prochilodus mariae). Revista UNELLEZ de Ciencia y Tecnología. (Guanare, Ven.). Serie: Producción Agrícola Año 1 (1 ): 11. 1983. 9. LOPEZ, H. y A. MACHADO-ALLISON. Algunos aspectos del desarrollo y crecimiento de Loricaria laticeps (Osteichtyes, Siluriformes, Loricaridae). Acta Biol. Venez. 9: 51-76. 1975. 10. MACHADO-ALLISON, A. Etapas del desarrollo del pez Lebiasina erythrinoides (Reganz, 1903). Acta Biol. Venez. 8: 579-622. 1974. 1 11. MACHADO-ALLISON, A. y H. LOPEZ. Etapas del desarrollo de Loricaricthyes tipus (Bleeker, 1864) (Osteicthyes, Siluriformes, Lori- caridae). Acta Biol. Venez. 9: 93-119. 1975. 12. MACHADO-ALLISON, A. Estudios sobre la subfamilia Serrasalminae (Teleostii: Characidae). Parte I. Estudio comparado de los juveniles de "Cachamas" de Venezuela. Acta. Biol. Venez. 2: 1-102. 1982. 13. MACHADO-ALLISON, A. y ZARRET. Datos sobre la biología reproductiva de Hoplosternum littorale de Venezuela. Acta Cient. Venezolana 35: 142-146. 1984. 14. MACHADO-ALLISON, A. Aspectos sobre la historia natural del "Cu rito" Hoplosternum littorale (Hancock, 1828) en el bajo Llano de Venezuela: desarrollo, alimentación y distribución espacial. Acta Cient. Venezolana 37: 72- 78. 1986. 15. PARKER, G. The optic quiasma in teleosts and its bearing on the asymmetry of the Heterostomata (flat fishes) .Bulletin M useum of Comparative Zoology Harvard College. Vol. 11, No.51903. 23 p. 16. PIRLOT, P. Morfología evolutiva de los cordados. Barcelona, España. Ediciones Omega. 1976. 966 p. 17. SALTHE, S. and W. E.DUELLMAN. Quantitative constrints associates with reproductive mode in anurans. ln: Vial, J. L. (Ed). Evolutionary biology of the anurans. University of Missouri Press. 1973. 470 p. 18. SETTE, S. Caracterización anatomo-fisiológica de tres especies de peces de la Cuenca Alta del río Uribante en relación a su fisiología reproductiva. Tesis de grado. Mérida, Ven. Fac. de Ciencias, Universidad de los Andes. 1986. 288p. 19. STEWART, R. La biología del bagre rayado Pseudoplatystoma fasciatum y Ptigrinum en la cuenca del río Apure, Venezuela. Revista UNELLEZ de Ciencia y Tecnología. Serie: Producción Agrícola (Guanare, Ven.) Año 1 (1): 13-41. 1983. 20. T APHORN, D. y G. LlL YESTROM. La relación largo, peso, fecundidad y dimorfismo sexual de Curimatus dalenae de la Cuenca del Lago de Maracaib6, Venezuela. Revista UNELLEZ de Ciencia y Tecnología (Guanare, Ven.) Año 1 (1): 73-78 p. 1983. 21. VENEZUELA. FONDO NACIONAL DE INVESTIGACIONES AGROPECUARIAs. Informe Anual 1985. Estación Experimental Mérida. Mérida, Ven. 1986. p. 130-140.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||