|
RESUMEN
Se realizó el
estudio del estómago e intestino grueso del báquiro de collar (Tayassu
tajacu), a través de muestras fijadas en formol al 10%. El estómago
presenta tres compartimientos o cámaras: lateral izquierda,
intermedia y lateral derecha. La cámara lateral izquierda presenta
dos divertículos, la intermedia presenta la desembocadura del
esófago y la derecha posee caracteres similares al estómago de los
monogástricos. Internamente el estómago está subdividido en tres
cámaras a través de pliegues. La cámara lateral izquierda presenta
mucosa glandular mucosecretora en su mayor extensión, en comparación
con la mucosa aglandular. El alimento es finamente reducido en esta
cámara, donde los olores y la estructura son compatibles con
procesos fermentativos. La cámara intermedia posee mucosa aglandular
en su mayor extensión, excepto en el área ventral opuesta al cardias
donde la mucosa es glandular, similar a la de la mucosa de la cámara
izquierda. Existe relación de contigüidad entre las mucosas
glandulares a través de islotes glandulares ventrales, siendo de
mayor desarrollo hacia la lateral izquierda, donde incluso se llega
en algunos casos a establecer una relación de continuidad a través
de una banda de mucosa glandular. La cámara lateral derecha presenta
extensiones de mucosa aglandular y áreas glandulares típicas del
estómago de monogástricos. El intestino grueso es muy desarrollado,
presenta saculaciones, apéndice cecal bien desarrollado y porción
ascendente del colon en forma de espiral, características típicas de
fermentador cecocólico. El estómago y el intestino grueso del
báquiro muestran caracteres morfológicos de adaptación a dietas
herbívoras, en especial a transición de frutívoro a folívoro. El
báquiro posee un ciego en general poco desarrollado, donde se
destaca un apéndice cecal que la mayoría de las veces es muy
prominente y por otra parte gran desarrollo del colon donde
destacan, la disposición en espiral del colon ascendente y la
presencia de saculaciones.
Palabras Clave: Báquiro; estómago; intestino grueso; macro.
STOMACH AND LARGE
INTESTINE MORPHOLOGY OF COLLARED PECCARY
(Tayassu tajacu)
SUMMARY
Stomach and large
intestine’s study of the collared peccari (Tayassu tajacu), was
carried out through samples fixed in 10% formalin. The stomach
presents three compartments or cameras: lateral left, intermediate
and lateral right. The camera lateral left presents two blind sacs,
the intermediate one presents the outlet of the esophagus and the
right possesses similar characters to the stomach of the
monogastrics. Internally, the stomach is subdivided in three cameras
through pleats. The lateral left camera presents glandular
mucosecreting mucous in its biggest extension, in comparison with
the aglandular mucous. The food is finely reduced in this camera,
where the scents and the structure are compatible with fermentative
process. The intermediate camera possesses mucous aglandular in
their biggest extension, except in the opposed ventral area to the
cardiac where the mucous one is glandular, similar to that of the
mucous of the left camera. Contiguous relationship exists among the
mucous glandular through ventral glandular islands, being of more
development toward the lateral left where you even ends up in some
cases to establish a relationship of continuity through a band of
mucous glandular. Lateral right camera presents extensions of mucous
aglandular and typical glandular areas of the monogastrics stomach.
The large intestine is very developed; it presents saculations,
appendix cecal developed and upward portion of the colon in hair
spring form, characteristic typical of cecocolic fermentator. The
stomach and large intestine of peccaries show morphology characters
of adaptation to herbivore diets, especially to frugivorous
transition to folivorous.
Keywords: Collared peccary; stomach; large intestine; gross.
INTRODUCCIÓN
La explotación zootécnica de animales de fauna silvestre con fines
productivos en nuestro país, ofrece grandes perspectivas de
desarrollo para el sub-sector pecuario nacional. En varias
oportunidades se ha planteado el estudio de especies de la fauna
silvestre, las cuales podrían explotarse racionalmente como fuente
de proteínas de alto valor biológico (Trebbau, 1970; González, 1970;
García, 1984; García, 1989).
En nuestro país, Trebbau (1970) planteó el estudio de animales de
fauna silvestre a través de los Parques Zoológicos. El alto costo de
las materias primas e insumos agrícolas lleva a buscar otras fuentes
proteicas alternativas más económicas que las tradicionales, y es
aquí donde toma vigencia la búsqueda de estos recursos en nuestra
fauna silvestre (chigüires, lapas, picures, báquiros, entre otros).
Es de suma importancia, el conocimiento de las características
morfológicas de animales de fauna silvestre, ya que servirá de base
a los estudios fisiológicos, fisiopatológicos, nutricionales,
reproductivos y patológicos, entre otros, lo cual unido al
conocimiento de su conducta y hábitos ecológicos, son aspectos de
gran interés a considerar para la posible explotación racional de
estas especies animales. La explotación racional de especies de
fauna silvestre, debe considerar dos aspectos fundamentales: la
producción de proteínas de origen animal para consumo humano a bajo
costo, y la conservación del equilibrio ecológico.
El báquiro, Tayassu tajacu, es una especie de la fauna silvestre que
se localiza desde el sur de Estados Unidos hasta Argentina. Presenta
una excelente adaptación a nuestro medio, de conducta alimentaria
poco exigente (forraje, raíces, frutas), y asociado a su potencial
cría en cautiverio, carne apetitosa y características reproductivas
que permiten su multiplicación en cautiverio, le conceden gran
potencial zootécnico. Sus condiciones biológicas llaman la atención
hacia el desarrollo de estudios integrales de esta especie, bajo
condiciones de cautiverio, en áreas tales como: morfología,
fisiología, patología, nutrición, reproducción, entre otros, así
como también hacia el incentivo de su utilización en la dieta del
campesino en un marco de equilibrio ecológico, que no ponga en
peligro la especie.
El báquiro es un mamífero del orden Artiodáctila, Familia
Tayassuidae que vive en rebaños y son vegetarianos (Fernández y
Ulloa, 1969; Fernández, 1970), llegándosele a considerar como animal
nocivo de cultivos tales como: yuca, ocumo, forrajes, frutas, entre
otros, cuando existen poblaciones elevadas de ellos (Fernández y
Ulloa, 1969). Habita en llanos y montañas, y se reproduce todo el
año montañas y se reproduce con una gestación de 142-148 días y con
2-4 crías por parto (Fernández y Ulloa, 1969; Argos, 1972).
En relación al tubo digestivo, Langer (1979) trabajó sobre el
significado del proventrículo del Pécari de collar; Tamate y Yamada (1983)
realizaron un aporte sobre la Histología e Inmunocitoquímica del
estómago posterior de la misma especie. En nuestro país existen
aportes anatómicos de la especie, tales como: morfología general de
la especie (Arias, 1986), donde se plantea la necesidad de realizar
estudios histológicos e histoquímicos del estómago e intestinos del
pécari de collar. También se han realizado trabajos en irrigación
arterial de los aparatos: genital masculino (Bendicho, 1994),
genital femenino (Pradere, 1991) y urinario (Campos, 1992).
En relación al estómago del báquiro, Tamate y Yamada, (1983)
refieren la presencia de cuatro cámaras, describiendo en el estómago
posterior la presencia de una mucosa pilórica, glándulas cardíacas y
glándulas fúndicas, con evidencia inmunocitoquímica de tres clases
de células endocrinas; Arias (1986) refiere la presencia de un
estómago con dos grandes cavidades, cada una con regiones
glandulares y aglandulares; Domínguez-Bello et al. (1989) describen
al estómago como voluminoso y complejo, constituido por tres cámaras
y seis tipos topográficamente diferentes de epitelios. En general,
el estómago está conformado por cuatro regiones: esofágica, cardial,
fúndica y pilórica.
Región Esofágica (parte de proventrículos). Se corresponde con el
área que continúa al esófago. Se caracteriza por poseer una mucosa
aglandular, revestida por epitelio plano estratificado, cuyo máximo
desarrollo lo presentan los rumiantes, conformando sus
proventrículos (rumen, retículo y omaso), pero puede estar ausente
en otras especies (Stinson y Calhoun, 1993; Banks, 1993).
Región Cardial (parte de las glándulas cardiales). Corresponde al
inicio de la región glandular del estómago. Se caracteriza por
presentar epitelio cilíndrico simple mucosecretor, que se invagina y
forma las glándulas cardiales tubulosas simples ramificadas, cuyo
principal componente corresponde a las células mucosecretoras, aún
cuando también existen células enteroendocrinas. Las células de las
glándulas son cilíndricas bajas o cúbicas; pueden alternarse con
células parietales o principales de acuerdo a la especie (Stinson y
Calhoun, 1993; Banks, 1993).
Región Fúndica (parte de las glándulas gástricas). También presenta
epitelio cilíndrico mucosecretor. Se invagina formando glándulas
tubulosas simples, que corresponden a las glándulas fúndicas, las
cuales son más largas, rectas y numerosas, en comparación con las
glándulas de la región precedente. Los componentes celulares de la
región son: células principales o cimógenas productoras de
pepsinógeno, pro-renina y lipasa gástrica; células parietales
productoras de ácido clorhídrico y factor intrínseco; células
mucosas del cuello, y células enteroendocrinas responsables de la
producción de serotonina gástrica, secretina y otras (Stinson y
Calhoun, 1993; Banks, 1993).
Región Pilórica. El epitelio de revestimiento es similar al descrito
en las regiones antes citadas, con corion de glándulas tubulosas
simples ramificadas, cuyo elemento celular predominante corresponde
a células mucosecretoras. Presenta también células enteroendocrinas
(Stinson y Calhoun, 1993; Banks, 1993; García, 1989). El báquiro
posee un ciego en general poco desarrollado, donde se destaca un
apéndice cecal que la mayoría de las veces es muy prominente.
Por lo antes expuesto, la investigación reseñada en este trabajo
estuvo orientada al estudio de los siguientes objetivos: aportar
información sobre la conformación del estómago e intestino grueso
del báquiro de collar (Tayassu tajacu), esto dentro del marco de un
estudio mayor que abarcó la descripción histológica, histoquímica y
ultraestructural del tubo digestivo (García, 1993).
MATERIALES Y MÉTODOS
Se procesaron muestras de quince (15) estómagos de báquiros de
collar (Tayassu tajacu) provenientes de una sobre población del
Zoológico Las Delicias, Maracay, estado Aragua. La muerte de los
animales se produjo por traumatismo craneano.
Previa identificación de las muestras, se fijaron en formol neutro
tamponado al 10%.
RESULTADOS Y DISCUSIÓN
Estómago
Estructura externa
Es un órgano con una estructura voluminosa y compleja, en la que se
pueden describir externamente tres cámaras o compartimientos,
fácilmente identificables: cámara lateral izquierda, cámara
esofágica o compartimiento intermedio, y cámara lateral derecha o
estómago verdadero (Figura 1).
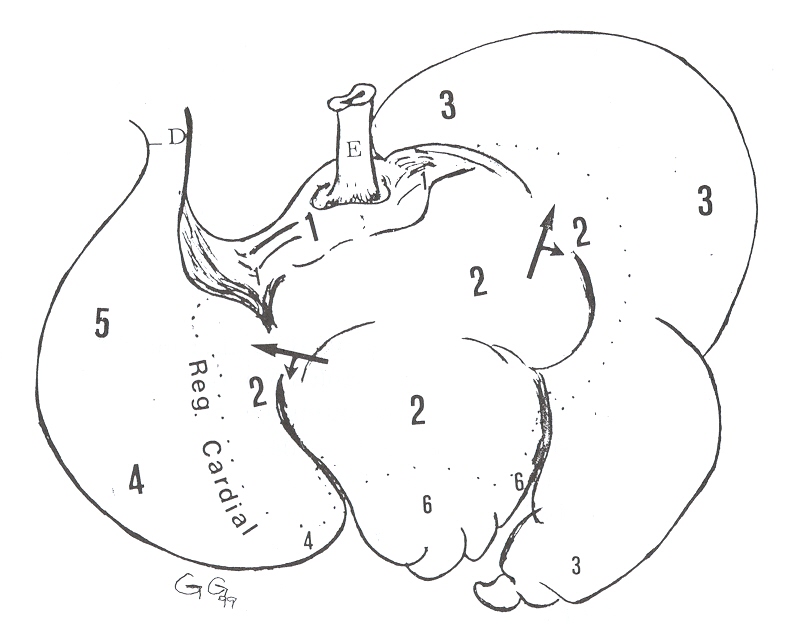 |
|
FIGURA 1. Estómago de báquiro, vista
dorso-lateral. E= esófago; D= duodeno; 1= surco o gotera; 2= región
esofágica; 3= región glandular (compartimiento ll); 4= región
fúndica; 5= región pilórica; 6= región glandular (compartimiento
intermedio). |
Estructura interna
Compartimiento lateral izquierdo: como su nombre lo indica, está
ubicada a la izquierda del plano medio. Se caracteriza por presentar
dos sacos o divertículos unidos entre sí, por un área algo tubular.
Macroscópicamente presenta una mucosa de aspecto rosado pálido y
paredes muy delgadas. La mucosa (se la denominó en este trabajo como
región 3) presenta pliegues cortos, gruesos, de dirección
predominantemente transversal, pero con pliegues longitudinales y
oblicuos que se entrecruzan. Es glandular, excepto en las cercanías
de los pliegues que conforman el orificio que une a esta cámara con
la intermedia, donde se evidencia claramente los límites de la
mucosa glandular con la mucosa aglandular de la cámara esofágica
(intermedia); sin embargo, se continúa hacia la cámara esofágica en
algunos animales a través de una estrecha banda, pero en otros, lo
hace a través de una cadena de islotes de mucosa glandular que se
localizan en el área que ocuparía la mencionada banda, hacia la
región ventral, opuesta al cardias, que adopta un aspecto ovalado
(se denominó región 6), de desarrollo variable con el animal
observado, la cual se describirá posteriormente (Figuras 1 y 2).
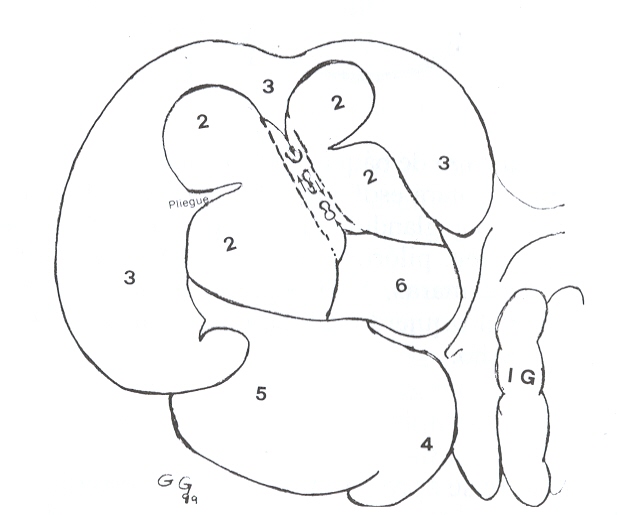 |
|
FIGURA 2. Estómago de báquiro, vista caudal. 2= región esofágica
(compart.
l); 3= región glandular (compart.II ); 4= reg. fúndica; 5= reg.
pilórica; 6= reg. gland. compart. I; flechas= islote o banda que une
mucosas 3 y 6. IG= intest. grueso. |
Compartimiento
intermedio: En este compartimiento desemboca el esófago, y se
encuentra parcialmente separado del compartimiento lateral
izquierdo, por pliegues que delimitan un orificio inter-cámaras, y
por otra parte, está también parcialmente separada del estómago
verdadero o cámara lateral derecha, por pliegues más bajos (de menor
desarrollo) en comparación con los pliegues primeramente mencionados
(Figura 3).
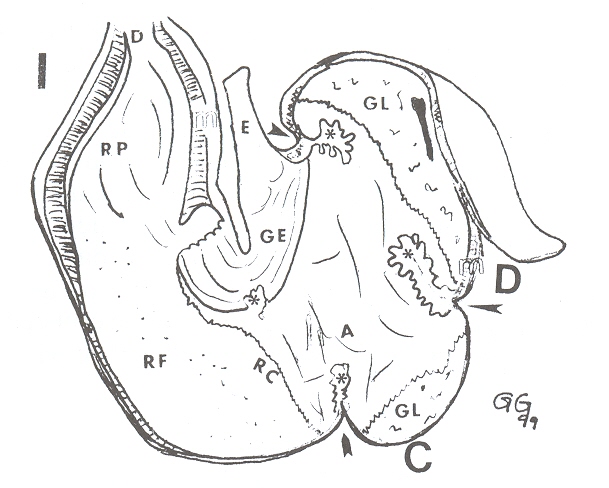 |
|
FIGURA 3. Estómago de báquiro, vista lateral
interna. E= esófago; GE= gotera esofágica; A= reg. aglandular
(esofágica); GL= reg. glandular; RC= reg. cardial; RF= reg.
fúndica; RP= reg. pilórica; m= capa muscular; flechas= surco
intercámaras; *=
pliegue intercámaras; I= compart. lateral izquierdo; C= comp.
intermedio; D= comp. lat. derecho. |
Macroscópicamente se aprecian tres regiones, como se expone en la
Figura 1:
Región 1: Caracterizada por presentar una mucosa de aspecto
blanquecino, aglandular, de pliegues longitudinales cortos, gruesos
y paralelos entre sí que recuerdan la gotera esofágica de los
rumiantes. Se extiende desde la desembocadura del esófago hacia la
izquierda del plano medio, hasta el pliegue que separa esta cámara
de la anterior, y hacia la derecha del plano medio hasta la región
pilórica del estómago verdadero (cámara lateral derecha). La pared
muscular a este nivel es muy desarrollada.
Región 2: Se caracteriza por presentar una mucosa grisácea, de
pliegues bajos y numerosos, de dirección predominantemente
transversal, con algunos longitudinales; rodea a la región 1,
excepto en la zona donde ésta se continúa con el estómago verdadero;
ocupa la mayor parte del área de la cámara esofágica, incluyendo la
de los pliegues que la separan de la cámara lateral izquierda. Sólo
existe un área de forma aproximadamente circular, de posición
ventral que es ocupada por la región 6.
Región 6: Se corresponde con la región esofágica de la mucosa
glandular de la cámara lateral izquierda, que como se mencionó
anteriormente, es un área glandular de aspecto generalmente
circular, opuesto al cardias, que puede prolongarse hacia la región
3 de la cámara lateral izquierda, bien sea a través de
prolongaciones a manera de islotes o bien a través de una banda
continua de mucosa, como se describió anteriormente. Existen islotes
glandulares, menos desarrollados en comparación a los citados, que
se extienden hacia la cámara lateral derecha.
Compartimiento lateral derecho o estómago verdadero: Este
compartimiento posee dos zonas claramente definidas de mucosa
glandular:
Región 4 o Región Fúndica: De aspecto liso, color gris-rosado, de
paredes muy delgadas, ocupa la curvatura mayor y región derecha del
estómago verdadero. Se relaciona con la mucosa aglandular de la
cámara esofágica, con límites claros de demarcación. Se torna pálida
hacia la región pilórica, región ésta con la que se continúa de
manera insensible.
Región 5 o Región Pilórica: Área de color blanquecino a rosado muy
pálida, de pliegues longitudinales gruesos, con mucosa glandular
lisa y paredes muy desarrolladas, especialmente hacia la cercanías
del duodeno donde forma el esfínter pilórico. Se extiende hacia la
derecha del plano medio donde forma el fondo pilórico. Intestino
grueso El intestino grueso del báquiro es muy desarrollado (Figura
4). Llama la atención la presencia de un apéndice cecal bien
desarrollado en la mayoría de las muestras observadas. También a
nivel del colon es importante resaltar la disposición en espiral del
colon ascendente y la presencia de saculaciones en el intestino
grueso.
El estómago presenta una estructura anatómica e histológica
compleja, formada por tres compartimientos, acorde con lo citado por
Domínguez-Bello et al. (1989), y por Campo (1992), pero se difiere
de lo descrito por Arias (1986), quien lo reporta como conformado
por dos compartimientos, así como también de lo señalado por Langer
(1979) y por Tamate y Yamada (1983), quiénes reportan la presencia
de cuatro compartimientos. De lo antes expuesto, se puede considerar
al estómago del báquiro, como intermedio entre el estómago de los
monogástricos y de los rumiantes, coincidiendo en cierta forma con
lo reportado por Langer (1974), cuando señaló que el estómago del
báquiro representa una forma intermedia entre el estómago sencillo
del cerdo y el altamente avanzado del hipopótamo, pero no llega a la
especialización adquirida por camellos y rumiantes.
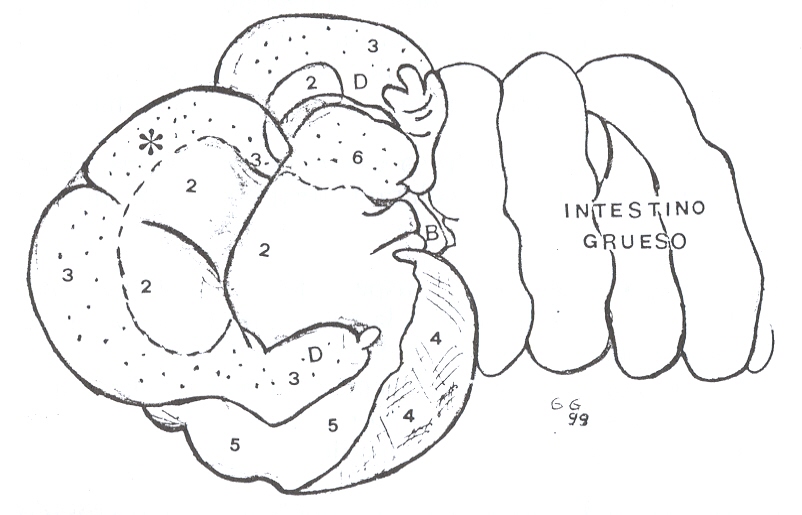 |
|
FIGURA 4. Estómago e intestino grueso de
báquiro. 2: Reg. esofágica; 3: región glandular mucosecretora; 4:
reg. fúndica; 5: reg. pilórica; 6: reg. glandular mucosecretora. *:
porción tubular que une a los divertículos; D: divertículos; B;
bazo. |
El estómago tiene gran importancia no solo funcional, sino también
permite observar la evolución taxonómica de las especies; y es así
como se dice que los mamíferos cecocólicos o fermentadores de
intestino grueso, representan una diferenciación más antigua que la
de los rumiantes (Langer y Snipes, 1991).
Anatómicamente el estómago consta de tres regiones, de acuerdo a la
pared muscular: Fornix Ventriculi, Corpus Ventriculi y Pars
Pilarica. Las mayores variaciones ocurren en el lado izquierdo del
fornix, y en menor número de especies existe variaciones del lado
derecho del fornix, decreciendo las diferenciaciones y
modificaciones hacia el cuerpo y región pilórica. De ahí que se
considere a las especies más evolucionadas a los rumiantes y a los
Tylópodas (camellos y llamas), y menos evolucionados a los no
rumiantes (suiformes), correspondiéndole a los hipopótamos una
escala mayor de desarrollo, al evolucionar en ellos tanto el lado
izquierdo como el derecho de la región fornical, así como la región
del cuerpo, en comparación con los Tayasuidos (báquiros), los cuales
presentan desarrollo evolutivo del lado izquierdo de la región
fornical, y la aparición de un pliegue entre el compartimiento
intermedio y el derecho en la región del cuerpo, acorde con lo
citado por Langer y Snipes (1991).
En la región fornical del báquiro aparecen dos divertículos y una
cámara intermedia (Langer, 1974), donde se observan epitelios tanto
glandulares como plano estratificado, similar a las especies
Camélidos (llamas y guanaco) e hipopótamos (Vallenas et al., 1971; Langer, 1974; Clemens y Maloiy, 1982; Domínguez-Bello
et al., 1989; Langer y Snipes, 1991). Se debe observar además, que el báquiro es
más evolucionado que el cerdo, el cual solo presenta un divertículo
glandular en la región fornical (Langer, 1974).
El incremento en el tamaño del estómago del báquiro, se traduce en
aumento de superficie (proyecciones digitiformes) en la mucosa
aglandular del compartimiento izquierdo, como se señala en este
trabajo, con presencia de pliegues que facilitan no solo los
procesos fermentativos, sino que también retardan el pasaje de la
digesta y permiten la selección de las partículas ingeridas de
acuerdo al tamaño, por medio de un efecto mecánico para su
transporte hacia la región posterior del estómago e intestinos, todo
lo cual está en concordancia con lo señalado por Langer y Snipes
(1991) y por Chivers y Hladik (1980), en relación a las adaptaciones
estructurales de los folívoros.
Aun cuando el estómago del báquiro no puede subdividirse de acuerdo
a la constitución histológica de la mucosa, tal como lo señalan
Langer y Snipes (1991), las regiones proventriculares, en general,
están delimitadas por epitelio cardíaco (cardial) o por epitelio
plano estratificado, pero puede haber la presencia de ambos tipos de
epitelio, como se describen en los resultados del presente trabajo,
y en los resultados obtenidos en llamas y guanaco por Vallenas et
al. (1971). Se les atribuye a estos tipos de epitelios funciones de
absorción, de transporte epitelial, y también de secreción de mucus
(Vallenas et al., 1971; Langer y Snipes, 1991).
La cámara intermedia es el lugar de desembocadura del esófago. Está
separada de las cámaras vecinas por pliegues que delimitan
orificios. Macroscópicamente e histológicamente se observan tres
áreas o zonas de mucosa:
a) El área contigua al esófago, blanquecina, de pliegues gruesos
paralelos entre sí, que siguen el sentido de la curvatura menor del
estómago, formando un surco o gotera esofágica similar a la de los
rumiantes, tal como lo señalan Arias (1986) y Domínguez-Bello et al.
(1989), con un epitelio plano estratificado de gruesa capa córnea,
similar al epitelio del esófago, y muscular de la mucosa muy
desarrollada.
b) Excepto en la zona contigua a la región pilórica, está rodeada
por una mucosa de color grisáceo, de escasos pliegues, que ocupa la
mayor parte de la cámara, que acorde con lo reportado por
Domínguez-Bello et al. (1989), corresponde aproximadamente al 83%
del compartimiento; se extiende corto trecho hacia las cámaras
vecinas, al formar y sobrepasar los pliegues intercámaras (Figura
1). Está formada por epitelio plano estratificado, con capa córnea
menos desarrollada al compararla con la región del surco,
correspondiendo a la región esofágica del estómago; la muscular de
la mucosa es poco desarrollada, siendo sustituida por una delgada
banda de tejido conjuntivo, en la medida que el corte se efectúa
lejos del área próxima al surco.
c) Una tercera mucosa se observa en el fondo de esta cámara
intermedia, de color rosado y pliegues cortos; está delimitada de la
región esofágica por un borde blanquecino, que a su vez se
corresponde con un incremento de la capa córnea en la zona de
transición epitelial. En esta región se observa un epitelio
cilíndrico mucosecretor y glándulas tubulosas, de células de
contenido citoplasmático granular eosinofílico. Esta área
corresponde al 16,72 % aproximadamente, de la cámara intermedia o
cámara esofágica, como se desprende de lo reportado por
Domínguez-Bello et al. (1989).
Vale la pena señalar que existe cierta proyección de la mucosa
glandular hacia la cámara lateral izquierda, llegando en algunos
casos a existir una verdadera relación de continuidad, y otra
proyección menos evidente hacia la cámara lateral derecha (Figura
2). De acuerdo a lo antes señalado, se puede denominar a esta cámara
intermedia como cámara esofágica, en lugar de “cámara cardial” como
lo hacen Domínguez-Bello et al. (1989), por ser la mucosa esofágica
el factor o área predominante, sobre todo si consideramos que el
área glandular correspondiente es relativamente poco desarrollada.
En cuanto a la cámara lateral izquierda, posee dos divertículos
unidos entre si por un área aproximadamente tubular (Figura 4). Está
conformada principalmente por una mucosa de epitelio cilíndrico
mucosecretor y glándulas tubulosas con células de contenido granular
eosinofílico (García, 1993). Esta área trata de continuarse con el
área glandular de la cámara intermedia, lo cual como se señaló
antes, logra en algunos casos a través de una banda estrecha (Figura
2), pero que cuando ésta no está presente, se observa una serie de
estructuras irregulares de mucosa similares a la mucosa glandular,
siendo la estructura histológica también semejante, encontrándose
bien delimitada del área esofágica por el borde blanquecino, como lo
hace con el resto del área glandular de la cámara lateral izquierda.
Se debe recordar que existe una pequeña área de epitelio esofágico
que se ubica en el inicio de la cámara lateral, en las proximidades
del pliegue intercámaras o pliegue fornical, tal como lo señalan
Domínguez-Bello et al., 1989 (Figuras 2 y 3).
La cámara lateral
derecha se corresponde con el estómago de los monogástricos, donde
se describen las regiones típicas del estómago: región esofágica
(región 2), representada por una estrecha no grueso, y fermentadores
preestomacales cuando el proceso se verifica en las expansiones
preestomacales (proventrículos), siendo filogenéticamente más
evolucionados los últimos, especialmente los rumiantes (Langer y
Snipes, 1991).
Los animales zoófagos puros, cuando poseen ciego, este es pequeño en
relación a todo el intestino grueso; además, es poco diferenciado al
igual que el colon. En cambio, los fitófagos poseen ciego y colon
bien desarrollados (Langer y Snipes, 1991). El volumen cecal
generalmente se incrementa en relación inversa a la calidad del
alimento (Langer y Snipes, 1991), pero existen casos excepcionales
como el hipopótamo que carece de esta región cecal (Clemens y Maloiy,
1982; MacDonald y Hartman, 1983; Langer y Snipes, 1991), lo cual
puede estar en relación a la importancia de la fermentación pre-estomacal
de esta especie (Clemens y Maloiy, 1982; Langer y Snipes, 1991).
En el intestino grueso (Figura 4) llama la atención la presencia de
un apéndice cecal bien desarrollado, tal como lo señaló Arias
(1986), y la disposición en espiral del colon ascendente, así como
la presencia de saculaciones, similar a lo que sucede en rumiantes,
suinos y roedores como lapas (Agouti paca) y picures, Dasyprocta
rubrata (Stinson y Calhoun, 1993; Banks, 1993; García, 1984; García,
1989).
En general, el báquiro posee un ciego poco desarrollado, donde se
destaca un apéndice cecal que la mayoría de las veces es muy
prominente. Este hecho tiene significación desde dos puntos de
vista: filogenético y morfofuncional. En el primer caso pone en
evidencia su transición hacia otro suiforme filogenéticamente más
evolucionado, el hipopótamo, tal como lo señalan Langer y Snipes
(1991); y en el segundo caso, permite asociar este hecho, por una
parte con el desarrollo del estómago y su papel en procesos
fermentativos, tal como se ha sugerido en hipopótamos (Clemens y
Maloiy, 1982; Langer y Snipes, 1991), y lo cual también sucede en
báquiros, y por otra parte con el gran desarrollo del colon (típico
de fermentadores de intestino grueso), donde destacan la disposición
en espiral de parte del colon ascendente y la presencia de
saculaciones. Se debe considerar que en los herbívoros, el ciego
incrementa su volumen con dietas de baja calidad, pero en este caso,
hay procesos digestivos previos en los compartimientos estomacales,
que en cierta forma mejoran la digestibilidad de la ingesta, y al no
haber en el báquiro incremento de volumen del ciego, no se presenta
el concomitante decremento del volumen relativo del colon. Esto está
acorde con lo descrito por Langer y Snipes (1991), para
fermentadores de intestino grueso (cecocólicos).
De lo antes descrito, se concluye que las características del
estómago e intestino grueso del báquiro, muestran caracteres
morfológicos de adaptación a dietas herbívoras, observándose de
manera más evidente, la transición alimenticia de frugívoros a
folívoros, coincidiendo con lo citado por Chivers y Hladik (1980),
cuando refieren que hay realmente una continuación de dieta de
frugívoros a folívoros en cerdos y pecaris.
CONCLUSIÓN
El estómago voluminoso y complejo, representa una transición,
entre el estómago de poligástricos y de monogástricos. El intestino
grueso presenta gran desarrollo, lo cual permite ubicarlo en
posición intermedia entre los fermentedores cecocólicos típicos y
los fermentadores pre-estomacales. Las características
morfofuncionales del tubo digestivo del báquiro (estómago-intestino grueso), permiten una mejor utilización de la ingesta, aun
en caso de dietas de baja calidad. Esto, conjuntamente con su
reproducción en cautiverio, implican la posibilidad real de
explotación intensiva o semi-intensiva de la especie, como fuente
alternativa de proteína de origen animal para consumo humano.
AGRADECIMIENTO
Al Consejo de Desarrollo Científico y Humanístico de la Universidad
Central de Venezuela (UCV), por financiar parcialmente este trabajo.
A la Universidad Central de Venezuela, Laboratorio de Histoquímica,
Cátedra de Histología, Facultad de Ciencias Veterinarias, por las
facilidades para desarrollar la investigación.
BIBLIOGRAFÍA
ARIAS, G. de FERNÁNDEZ. 1986. Anatomía Regional del Pécari de Collar
(Tayassu tajacu, Linneo) Comparada con el Cerdo Doméstico. Trabajo
de Ascenso a la categoría de Titular. Maracay. Universidad Central
de Venezuela. Facultad de Ciencias Veterinarias. 241 p.
BANKS, W. 1993. Applied Veterinary Histology. Williams y Wilkins.
Baltimore. pp. 373-423.
BENDICHO M., M. 1994. Irrigación arterial del Aparato Genital
Masculino del pécari de collar (Tayasu tajacu). Trabajo de Ascenso
para la categoría de Asociado. Maracay. Universidad Central de
Venezuela. Facultad de Ciencias Veterinarias. 72 p.
CAMPO R., G. 1992. Arteria Celíaca del Pécari de Collar (Tayassu
tajacu). Trabajo de Ascenso para la categoría de Asociado. Maracay,
Universidad Central de Venezuela. Facultad de Ciencias Veterinarias.
78 p.
CHIVERS, D. J. and C. M. HLADIK. 1980. Morphology of the
Gastrointestinal Tracts in Primates: Comparations with others
Mammals in Relation with Diet. J. Morph. 166:337-386.
CLEMENS, E. T. and G. M. O. MALOIY. 1982. The Digestive Physiology
of Three East African Herbivores: The Elephant, Rhiniceros and
Hippopotamus. J. Zool. 198:141-156.
DOMÍNGUEZ-BELLO, G., R. PARRA y A. BALDIZAN. 1989. Características
Digestivas del Báquiro de Collar (Tayassu tajacu). Resultados
Preliminares. Acta Científica Venezolana. 40:381-384.
FERNÁNDEZ YÉPEZ, A. y G. ULLOA. 1969. Mamíferos de Venezuela.
Maracay. Universidad Central de Venezuela. 102 p.
FERNÁNDEZ YÉPEZ, A. y G. ULLOA. 1970. Claves para la Determinación
de Ordenes, Familias y Géneros de Mamíferos en Venezuela. Maracay.
Universidad Central de Venezuela. Facultad de Agronomía. 65 p.
GARCÍA C., G. 1984. Aspectos Morfológicos e Histoquímicos del Tubo
Digestivo de la Lapa (Agouti paca). Trabajo de Ascenso a la
categoría de Asistente. Maracay. Universidad Central de Venezuela.
Facultad de Ciencias Veterinarias. 80 p.
GARCÍA C., G. 1989. Histología e Histoquímica del tubo Digestivo del
Picure (Dasyprocta rubrata). Veterinaria Trop. 14:53-83.
GARCÍA C., G. 1993. Aportes Histógicos, Histoquímicos y
Ultraestructurales, sobre el tubo digestivo del báquiro de collar (Tayassu
tajacu). Trabajo especial de grado para obtener el título de
Magister Scientiarum. Maracay. Universidad Central de Venezuela.
Facultad de Ciencias Veterinarias.
GONZÁLEZ G., E. 1970. Investigación Biológica y Fauna Silvestre.
Defensa de la Naturaleza. Caracas. 1(1):34-37.
LANGER, P. 1979. Adaptational Significance of the Forestomach of the
Collered Peccary, Dicotyles tajacu (L. 1758) (Mammalia: Artiodactyla).
Mammalia 43:235-245.
LANGER, P. and R. SNIPES. 1991. Adaptations of Gut Structure to The
Function in Herbivores. In: Tsuda T., Sasaki Y., Kawashima R. (EDS.).
Physiological Aspects of Digestion and Metabolism in Ruminants.
Academic Press. San Diego. USA. pp.349-384.
MacDONALD, A. and W. HARTMAN. 1983. Comparative and Functional
Morphology of the Stomach in the Adult and Newborn Pigmy
Hippopotamus (Cheoropsis liberiensis). J. Morpho. 177:269-276.
PRADERE G., J. 1991. Anatomía Vascular del Aparato Genital Femenino
del Pécari de Collar (Tayassu tajacu). Trabajo de Ascenso a la
Categoría de profesor Asociado. Maracay. Universidad Central de
Venezuela. Facultad de Ciencias Veterinarias. 63 p.
SCHMIDT-NIELSEN, N. 1976. Fisiología Animal: Adaptación y medio
ambiente. Editorial Omega S.A., Barcelona, España. pp.121-138.
STINSON, A. W. y M. L. CALHOUN. 1993. Sistema Digestivo.
In: Dellmann y Brown, eds. Histología Veterinaria. Edit. Acribia.
Zaragoza, España. pp. 223-283.
TAMATE, H. and J. YAMADA. 1983. Histological and Inmunocitochemicals
observation of the Hindstomach of the Collared Peccary, Dycotiles
tajacu (Tayassuidae). Jpn. J. Vet. Sci. 45(5):547-559.
TREBBAU, P. 1970. Importancia de los Parques Zoológicos en la
Conservación de la Fauna. Defensa de la Naturaleza. Caracas.
1(1):17-19.
VALLENAS, A., J. F. CUMMINGS and R. MUNNEL. 1971. A Gross Study of
the Compartmentalized Stomach of two New-World Camelids. The Llama
and Guanaco. J. Morph. 134:399-424.
|