|
Zootecnia Trop.,13(1):31-48 1995 ASPECTOS REPRODUCTIVOS Y BIOMÉTRICOS DEL
PARGO ALETA NEGRA, Gabriel Gómez, G.; R. Guzmán; T. Ramírez y L. Marcano. FONAIAP. Centro de investigaciones
Agropecuarias el |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
INTRODUCCIÓN La familia Lutjanidae, a nivel mundial, está conformada por 23 géneros y cerca de 230 especies, Nelson (24). El género Lutjanus, dentro del cual se ubica la especie en estudio, constituye uno de sus principales representantes con 69 especies, Druzhinin (4), de las cuales 8 han sido reportadas para Venezuela y 11 en total para el Atlántico Centro Occidental (2,8). L. buccanella, (Figura 1) se caracteriza por ser una especie de la plataforma y/o del talud. Habita sobre fondos coralinos, rocosos o arenosos a profundidades que oscilan entre 80 y 150 m. Sin embargo, los Juveniles de esta especie suelen vivir en aguas más someras, a menudo entre 35 y 50 m (8). En Venezuela es muy abundante, en áreas de las Islas Los Hermanos (2,8,10). Esta especie, es parte importante de la economía pesquera de algunos países como Cuba, lo cual ha motivado a realizar estudios relacionados con su biología y dinámica poblacional (Espinoza y Pozo (6), Pozo et al (17) y Espinoza et al (7)). A pesar de su importancia comercial, en Venezuela, no existen trabajos relacionados con la biología y pesquería de esta especie. Por tales razones, se creyó conveniente realizar el presente estudio como una contribución al conocimiento de aspectos importantes de su biología, tales como reproducción y estructura poblacional, entre otros. Estos parámetros son de utilidad para la interpretación del comportamiento de las poblaciones de peces ante ciertos niveles de explotación, y además sirven de base para la implementación de políticas pesqueras en función de un mejor aprovechamiento de los recursos pesqueros.
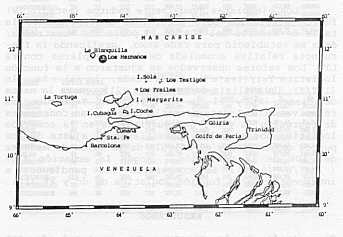
MATERIALES Y MÉTODOS Las muestras utilizadas en el presente estudio, se obtuvieron durante el período enero a diciembre de 1992, a través de campañas de pescas exploratorias en áreas de las Islas Los Hermanos, ubicadas al Noreste de la Isla de Margarita entre los 11°40' LN y los 64°23' LO (Figura 2). Las capturas se realizaron abordo de la embarcación "Golfo de Cariaco" del FONAIAP, utilizando nasas tipo antillanas. A los ejemplares capturados se les determinó la longitud total (mm) y el peso (g).
Esta información se utilizó para establecer la estructura de tallas de la población muestreada en base a un histograma de fecuencias, con intervalos de 20 mm y además se relaciono la longitud total y el peso mediante regresión lineal (21) para hembras y machos respectivamente; de acuerdo al ajuste de la curva de crecimiento relativo P = a*LTb, indicada por Ricker (18) y su expresión logarítmica: log (P) = log (a) + b * log (LT); en donde "a" y "b" son constantes de regresión. Ambas ecuaciones se compararon a través de un análisis de covarianza (20). Una prueba t-student (21), permtió establecer el tipo de crecimiento relativo en peso de la especie. Las gónadas fueron examinadas macroscópicamente, lo que permitió la separación de sexos. Cuya proporción se analizó por medio X2 (21) y permitió además establecer la madurez sexual, de acuerdo al criterio de Holden y Raitt (12), con lo cual se infirió sobre la época de actividad reproductiva de la especie. Los ejemplares sexualmente maduros (estadío IV) fueron utilizados para determinar analíticamente la talla de madurez del 50% de la población muestreada; ésta se estableció para cada sexo, graficando la frecuencia relativa acumulada de los ejemplares contra la longitud total. Los valores observados se ajustaron a la función logística Y = 1/(1+a*ebx), linealizada, ln (a) + bx = ln (1-Y/Y). Un análisis de regresión (21), entre la marca de clase (X) y el ln (1-Y/Y); con Y = frecuencias relativas acumuladas, permitió obtener las constantes de regresión (a y b), que sustituidas en la ecuación logística por su valor, permitieron el ajuste de los puntos observados y el cálculo de la talla de madurez del 50% de la población, según la ecuación X0,5 = (-lna)/b (9,22). Se compararon las pendientes e interceptos de la función logística de machos y hembras (20). RESULTADOS COMPOSICION DE LA MUESTRA: De un total de 855 ejemplares estudiados, 410 (47,95 %) fueron machos, con longitudes entre los 200 y 650 mm, y una talla media de 328 mm y 427 (49,94%) hembras, con tallas que oscilaron entre los 210 y 665 mm, cuya longitud promedio fue de 337 mm, Cuadro 1. En la muestra, se observaron además 18 (2,11%) ejemplares de sexo indeterminado, con longitudes comprendidas entre los 170 y 200 mm.
PROPORCION DE SEXOS: La relación machos : hembras por mes, se presenta en el Cuadro 2; se observa que durante el período analizado ésta no se alejó de la esperada (1:1), siendo la proporción total de 0,96:1 (P>0,05).
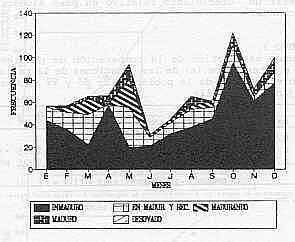
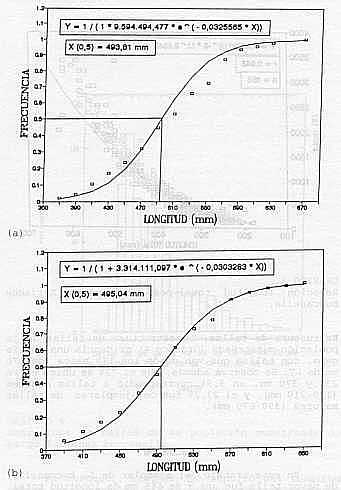
EPOCA DE REPRODUCCION: Los resultados indican la presencia de ejemplares en avanzado estado de madurez (estadío IV), durante los meses febrero - mayo, agosto y octubre - diciembre ; distinguiendose picos de mayor actividad reproductiva, en mayo, octubre y diciembre. Ejemplares desovados (estadío V) no fueron observados (Gráfico 1). TALLA DE MADUREZ DEL 50% DE LA POBLACION: La talla a la cual el 50% de la población es capaz de reproducirse, se estableció para los machos en 495 mm de longitud total, observándose ejemplares maduros a partir de los 390 mm. Para las hembras, esta talla se ubicó en 494 mm encontrándose individuos maduros desde los 370 mm de longitud total.
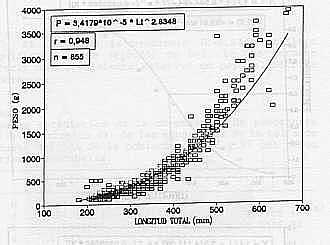
Las pruebas de pendientes (b) de las curvas de madurez, no mostraron diferencias significativas entre sí (p>0,05), lo contrario ocurrió para los interceptos, Cuadro 3. Por esta razón y a pesar de la similitud entre las tallas de madurez, se presentan dos curvas (Gráfico 2, a y b). RELACION LONGITUD - PESO: Las regresiones establecidas entre ambas variables para machos y hembras, respectivamente, no mostraron diferencias significativas (P>0,05), entre sexos, estimándose una ecuación común (Cuadro 4, Gráfico 3); que quedó expresada como:
El valor de la pendiente (b = 2,8348) difiere significativamente (P<0,05) de b = 3, lo que permite inferir un crecimiento relativo en peso alométrico minorante.
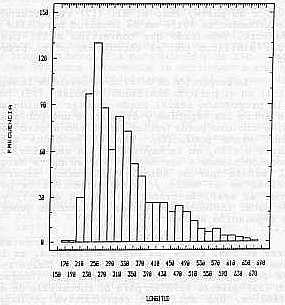
ESTRUCTURA DE TALLAS: La estructura de tallas de la población muestreada (Gráfico 4), evidencia una amplia gama, con tallas que van desde los 170 hasta los 670 mm de longitud total. Se observa además, que el 73% de la población se ubicó entre 230 y 370 mm, un 3,9 % correspondió a tallas menores; entre 170 y 210 mm y el 23,2 % fueron ejemplares de tallas mayores ubicadas entre los 390 y 670 mm de longitud total. DISCUSIÓN En este trabajo, el ejemplar de L. buccanella, de mayor talla fue una hembra de 665 mm de longitud total, mientras que el macho de mayor tamaño midió 650 mm. La literatura para esta especie registra tallas similares a las máximas encontradas en el presente estudio.
Por su parte, Pozo et al (17), registraron una talla promedio de 301,6 mm de longitud horquilla para la especie, resultado que convertido a longitud total, pudiera guardar similitud con el encontrado en el presente estudio. La proporción de machos:hembras no se desvió de la esperada (1:1) en el período analizado. Nikolsky (15) indica que la proporción sexual puede cambiar considerablemente entre las especies y diferir de una población a otra, e incluso puede variar de un año a otro dentro de una misma población; también menciona, que no obstante a ello en la mayoría de las especies, dicha proporción tiende a ser de 1:1. Por su parte Margalef (14), señala que en animales con sexos separados, dispersos en un medio y cuyos encuentros se pueden suponer al azar, la proporción sexual es 1:1; y que está influenciada además por cambios genéticos y de regulación ambiental. La presencia de ejemplares en avanzado estado de madurez en casi todos los meses analizados, es indicativa de un largo período reproductivo. Tales resultados concuerdan con los reportados por Thompson y Munro (23) quienes señalan para L. buccanella de aguas de Jamaica, una actividad reproductiva prolongada, indicando dos períodos, febrero - mayo y agosto - noviembre; y guardan relación con los encontrados por Pozo et al (17) en la plataforma Sur-oriental de Cuba, donde observaron ejemplares maduros y desovados de L. buccanella, desde marzo a octubre. Por otra parte, la ausencia de ejemplares desovados podría sugerir que el área de pesca no coincide con el área de reproducción de la especie; lo que posiblemente, pudiera estar relacionado con factores migratorios. La talla de madurez del 50% de la población, determinada para machos (495 mm LT) y para hembras (494 mm LT) mostró gran similitud. Estas tallas resultaron superiores a las determinadas por Pozo et al (17) para L. buccanella de Cuba, quienes señalaron una talla media de madurez de 210 - 250 mm de longitud horquilla para las hembras y de 260 - 330 mm para los machos y a las señaladas por Thompson y Munro (23) en aguas de Jamaica, quienes señalaron una talla media de madurez entre 250 - 270 mm de longitud horquilla para los machos, y de 230 - 250 mm, para las hembras; esta diferencia la atribuyen a una tasa de crecimiento más rápida en los machos, o que estos maduran más tardíamente que las hembras. El valor de la pendiente; b= 2,83 en la ecuación común de longitud - peso de machos y hembras de L. buccanella indica un crecimiento relativo en peso alométrico minorante, (P<0,05). Pozo et al (17) en Cuba, para esta misma especie indicaron un valor de b = 3,03 en la relación longitud - peso. Valores del coeficiente "b" han sido señalados para otras especies de Lutjanus; al respecto, Gómez et al (11) indicaron un valor de b = 3,02 para L. vivanus de Venezuela, Rubio et al (19) señalaron un valor de b = 2,97 para L. sinagris de Cuba, y Pozo (16) para esta misma región, indicó valores de b = 2,59 y b = 2,53, respectivamente para hembras y machos de L. analis. Le Creen (13) señala que la relación longitud - peso caracteriza a una población y que dicha relación puede ser descrita matemáticamente y agrega, que el coeficiente de regresión "b" de la misma, además de diferir entre especies y habitat, también varía con la época del año. Por otra parte, Bowering (1) señaló que las posibles diferencias en el crecimiento de machos y hembras, se deben más a factores genéticos que determinan la fisiología y comportamiento del pez, que a los factores ambientales. La composición por tallas de la población muestreada, indica que la misma podría estar sometida a una actividad pesquera intensa, ello fundamentado en que más del 70 % de los ejemplares se ubicó entre 230 y 370 mm de longitud total, longitudes éstas, inferiores a la talla de madurez del 50% de la población. La incidencia de los regímenes de pesca en los cambios que puede sufrir una población de peces, ha sido señalada por Csirke (5), quien destaca efectos sobre la estructura de edades o tamaño de los individuos, así como, en su tasa de crecimiento y en su capacidad reproductiva. Añade además, que uno de los cambios más notorios es la reducción en la proporción de peces viejos; situación que se evidenció en la estructura de tallas establecida en este estudio, en la que se observa un predominio de tallas medias y muy pocos ejemplares de tallas mayores. CONCLUSIONES
RESUMEN Se estudiaron aspectos reproductivos y biométricos del pargo aleta negra Lutjanus buccanella. Se analizó un total de 855 ejemplares capturados con nasas antillanas, de enero a diciembre de 1992. La proporción total de sexos (0,96:1), no se alejó (P>0,05) de la esperada (1:1). La especie mostró una actividad reproductiva prolongada que se intensifica en mayo, octubre y diciembre. La talla de madurez sexual del 50% se determinó en 495 mm para machos y 494 mm para hembras. La relación longitud - peso entre hembras y machos, no mostró diferencias significativas (P>0,05); se estimó una ecuación común definida como: P = 3,4179*10-5 * LT2,8348, con un crecimiento relativo alométrico minorante (P<0,05). La estructura de la población mostró un amplio intervalo de tallas (170 - 670 mm de longitud total); donde más del 70% de la muestra se ubicó entre los 230 - 370 mm de longitud total.
BIOMETRICS AND REPRODUCTIVE ASPECTS OF BLACK FIN SNAPPER Lutjanus buccanella (CUVIER, 1828) FROM ISLAND LOS HERMANOS, EASTERN, VENEZUELA. SUMMARY Reproductive and biometric aspects of the black fin snapper, Lutjanus buccanella, were studied. A total of 855 individuals, collected with antillean traps between january and december 1992, were examined. Sex ratio was estimated 0,96:1, which did not deviate from the 1:1 ratio (P>0,05). The species showed a prolonged reproductive activity period, with a greater intensity during May, Octuber and December. Sexual maturity size at 50% was estimated at 495 mm for males and 494 mm TL for females. Length - weight relationship did not showed a significantly difference between males and females (P>0,05), thus a common equation was estimated as W = 3,4179 *10-5 * TL2,8348, with alometric relative growth (P<0,05). Size structure (TL) showed an ample range , (170 - 670 mm); in which than more of the 70% of sample ocupied the range 230 - 370 mm TL. Key Words: Lutjanus buccanella, Sex ratio, Reproductive activity, sexual maturity, Size Structure AGRADECIMIENTO Expresamos nuestro agradecimiento a los técnicos Ramón Chacón, Andrés Urbaneja y Germán Vizcaíno por su participación en las campañas de pesca exploratoría, así como en los muestreos abordo. Un reconocimiento muy especial a la tripulación de la embarcación "Golfo de Cariaco". BIBLIOGRAFÍA 1. BOWERING, W. Distribution, age and growth, and sexual maturity of with flounder Glyptocephalus cynoglossus, in new foundland waters. J. Fisheries Res. Board can., 33:1574-1584. 1976. 2. CERVIGON, F. Los peces marinos de Venezuela. Fundación La Salle de Ciencias Naturales. Tomo II. Caracas. Venezuela. 994 p. 1966. 3. CERVIGON, F. Y W. FISCHER. Catálogo de especies marinas de interés económico actual o potencial para América Latina. parte 1. Atlántico Centro y Sur Occidental. Roma FAO/UNDP. Sic/1372 p. 1979. 4. DRUZHININ, A. D. The range and biology of snappers (Fam, Lutjanidae). J. Icthiology. 10 (5): 717 -735. 1970. 5. CSIRKE, S. Introducción a la Dinámica de población de peces. FAO. Doc. Tec. Peca (192): 82. 1980. 6. ESPINOZA, L. Y E. POZO. Edad y Crecimiento del sesi Lutjanus buccanella (Cuvier, 1828) en la Plataforma Suroriental de Cuba. Rev. Cub. Inv. Pesq. 7 (1): 80. 1982. 7. ESPINOZA, L. ; E. POZO Y R. QUEVEDO. Edad y Crecimiento del sesí Lutjanus buccanella (Cuvier, 1828) en la Plataforma Nororiental de Cuba. Rev. Cub. Inv. Pesq. 9(1-2): 9-20. 1984. 8. FISCHER, W. (ED). Species identification sheets for fishery purposes, Western Central Atlantic (Fisching Area 31) FAO Rome, Vols., 1-7. 1978. 9. GAERTNER, D. Y F. LALOE. Etude biometrique de la taille a'premiere maturité sexuelle de Geryon maritae, Maning et Holthuis, 1981 du Sénegal. Oceanol. Acta, 9 (4): 479 -487. 1986. 10. GOMEZ, G.; A. URBANEJA; R. CHACON Y G. VIZCAINO. Algunos aspectos biológicos del pargo aleta negra, Lutjanus buccanella (Cuvier, 1828) (Pices: Lutjanidae), de las Islas Los Hermanos, Venezuela. AsoVAC. XLI Conv. Anual. Acta Científica. Vol. 42 (1): 293. 1991. 11. GOMEZ, G.; R. CHACON; A. URBANEJA Y G. VIZCAINO. Aspectos preliminares de la biología del pargo ojo amarillo Lutjanus vivanus (Cuvie, 1828) (Pices: Lutjanidae), del Sureste de Las Islas Los Hermanos, Venezuela. AsoVAC. XLII. Conv. Anual. Acta Científica. Vol. 43(1): 279. 1992. 12. HOLDEN, M. J. Y D.F. RAITT. Manual de Ciencia Pesquera. Parte 2. Métodos para invistigar los recursos y su aplicación. Doc. Tec. F.A.O. Pesca (115) Rev. 1:211. 1975. 13. LE CREEN, E.D. The length-weigth relationship and seasonal 14. MARGALEF, R. Ecología. Omega S.A. Barcelona (España). 591 p. 1977. 15. NIKOLSKY, G. The ecology of fishes. Acedemic Press. New York. 185-187 p. 1963.. 16. POZO, E. Edad y Crecimiento del pargo Lutjanus analis (Cuvier, 1828) en la Plataforma Nororiental de Cuba. Rev. Cub. Inv.- Pesq. 4(2): 1-24. 1979.: 17. POZO, E. ; L. ESPINOZA y M. GUARDIOLA. Aspectos prelimina-res de la biología del sesi Lutjanus buccanella (Cuvier, 1828) en la Plataforma Suroriental de Cuba. Rev. Cub. Inv. Pesq. 8(3): 1-28. 1983. 18. RICKER, W. E. Computation and interpretation of biological statistical fish populations. Bull. Fish. Res. Board. Can. (191): 382 p. 1975. 19. RUBIO, R.; P. SALAHANGE Y M. BETANCOURT. Relaciones de la edad con el largo, el peso y la fecundidad de la biajaiba Lutjanus sinagris de la Plataforma Suroriental de Cuba. Rev. Inv. Pesq. 10 (1-4): 78-90. 1985. 20. SNEDECOR, G. AND W. COCHRAN. Statistical Methods. The Iowa 21. SOKAL, R. AND. ROHLF. Biometry. Freeman and Co, San Francisco, 859 p. 1981. 22. SOMERTON, D. A. A computer technique for estimating the size of sexual madurity in crabs. Ca. J. Fish. Aquacti. Sci. 47: 1488-1494. 1980. 23. THOMPSON, R. AND J.L. MUNRO. The biology, ecology and bioecomics of the snapper. Lutjanidae in Caribean Coral Ref Fisheries Resources. ed. by J.L. Munro. ICLARM Studies and Reviews (1983) 6: 94-109. 1974. 24. NELSON, J.S. Fishes of the word. John Willy & Sons, New York,E.E.U.U. 416 p. 1976. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

|
|