|
Zootecnia Trop., 16(1):41-60. 1998 EFECTO DE LA FERTILIZACIÓN NITROGENADA
SOBRE LOS COMPONENTES Romero C., S. Alfonso, R. Medina y R. Flores FONAIAP-Estación Experimental Falcón. Coro,
estado Falcón. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
INTRODUCCIÓN Existen numerosos trabajos que reportan el efecto del nitrógeno sobre la producción final de biomasa en pasturas; lo que ha permitido establecer criterios generales de manejo sobre una biomasa acumulada, sin analizar la cinética de elaboración y/o pérdida de esa fitomasa en el pastizal. El estudio de los fenómenos morfogenéticos donde se describe la dinámica de aparición, alargamiento, duración de vida y senescencia de órganos, que forman la estructura del vegetal permite medir la acción de variables, como la fertilización nitrogenada (ligada a las condiciones edafoclimáticas) y establecer la dinámica del crecimiento, producción de biomasa total y aprovechable, que ayudan a definir normas de manejo racional. El presente trabajo se realizó con el objeto de determinar la dinámica del crecimiento del pasto estrella sometido a la fertilización nitrogenada, realizando una descripción de la cinética de elaboración de biomasa y su evolución a través del tiempo, mediante el estudio de los componentes morfologicos; considerando que en las especies estoloniferas 14 producción de biomasa es el resultado del crecimiento de dos conjuntos de órganos: hojas y estólones, en proporciones que 1varían fuertemente en función de la edad del rebrote y la acción conjugada de las variables del medio (3,11,12). MATERIALES Y MÉTODOS Ubicación El ensayo se realizó en las Colonias de Araurima zona de Bajo Tocuyo estado Falcón, en la Unidad Agroecológica 'E156, con las siguientes características: clima de bosque seco tropical, período de lluvias de 8 a 9 meses con distribución de tipo bimodal (abril-julio y septiembre-diciembre), con valores que oscilan entre 800 y 1200 mm/año. La temperatura varía entre 21"C y 26"C. (9) Protocolo experimental Para medir el crecimiento y la senescencia de material vegetal, se instalaron 3 transéctas por parcela, a lo largo de las cuales se identificaron diez plantas con un cordel de nylon. Modificación de metodología descrita por Davies (4) y Romero (11). -Crecimiento: Se midió la tasa de acumulación de
la materia seca; expresada a través de: número de hojas y elongación de estolones
producidos por día. Este crecimiento se midió a través de:
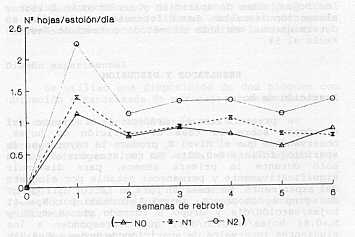
*Medición de crecimiento de tallos: En las plantas marcadas se midió la elongación de cada tallo o estolón, entre dos observaciones sucesivas. -Senescencia: Determinada semanalmente por la tasa de muerte o pérdida de materia seca viva en el pastizal, mediante la medición del número de hojas. En el mismo dispositivo utilizado para evaluar la aparición de hojas, se anotó el número de hojas que murió entre dos períodos de observación. El acumulado de hojas senescentes durante el ciclo de medición, reflejó el total de materia seca muerta. Estas observaciones permitieron además conocer la vida útil de las hojas. Diseño experimental Se utilizó una disposición de dos bloques con ubicación aleatorizada de parcelas o tratamientos. Tratamientos o factores estudiados: Nitrógeno: Tiempo: Variables estudiadas Tasa de aparición de hojas, número de hojas vivas (hojas verdes o parcialmente verdes), número de hojas muertas (senescencia foliar) , producción total de hojas, tasa de elongación de estolones y aparición de nuevas yemas o ramificaciones. Análisis y presentación de datos La información se analizó a través de programas de cálculo, una base de datos elaborada con el paquete computarizado Dbase III y análisis estadístico STATITCF. Aunque la unidad básica de medida es la yema vegetativa, es decir el punto de crecimiento de cada ramificación, los resultados se presentan por estolón, que constituye la planta entera, cuya expresión es más comprensible para efectos de la cuantificación del rendimiento. Se realizaron análisis de varianza (AV) para las variables hojas vivas, hojas muertas, total de hojas, tasa de aparición y senescencia foliar y elongación de tallos. Las diferencias entre medias se determinaron mediante el método o test de Newman-Keuls al 5%. RESULTADOS Y DISCUSIÓN Aparición de hojas Se presenta en el Gráfico 1 el efecto del nitrógeno sobre la tasa de aparición de hojas, observándose que el nivel N, produce la mayor tasa de aparición foliar (P<0,001) . Se registra una alta tasa solo durante la primera semana, para disminuir significativamente y permanecer estable por el resto del experimento. Se observan para el factor nitrógeno, dos grupos homogéneos. Grupo A formado por N,=1,41 hojas/estolón/día y Grupo B formado por N,=0,95 y N,=0,84 hojas/estolón/día, que corresponden a los siguientes intervalos de aparición de hojas: N,=0,71, N,=1,05 y N,,=1,19 días/hoja. Para el factor tiempo (semanas) los grupos se presentan en el Cuadro 2, indicando una tendencia o estrategia del pasto estrella a emitir rápidamente un número importante de hojas, independientemente del nivel de nitrógeno, solo que en N2 es mayor debido al efecto de este elemento sobre la tasa de elongación de estolones y aparición de nuevas yemas o ramificaciones (Gráficos 2 y 3) los cuales traen consigo la aparición de nuevas hojas. Estos resultados coinciden con los datos mostrados por Huguet (8), quien concluye que el ritmo de emisión foliar de un estolón es independiente del nivel de nutrición nitrogenada. Resultados similares presentó García (5) en mediciones, sobre pasto estrella sin fertilizar en la misma zona donde se realizó el presente trabajo, encontrando en 49 días de observación una tasa promedio de 0,76 hojas/estolón/día.
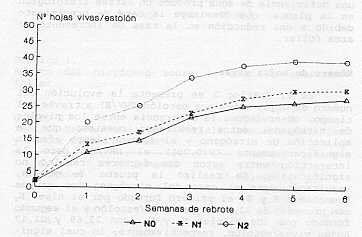
Es importante señalar que el efecto del nitrógeno sobre la dinámica del crecimiento (en este caso la tasa de aparición foliar) está afectado por las variables del medio, tales como la luminosidad. Varios autores citados por Anslow (1) señalan que esta tasa no es afectada por diferentes niveles de nitrógeno, pero existe una interacción entre alto nivel de este elemento y alta intensidad de iluminación, que producen un adecuado nivel de carbohidratos, necesario para originar un incremento significativo de la aparición foliar. La reducción del efecto nitrógeno es debida probablemente a pérdidas de este elemento y/o interacción con la disponibilidad hídrica del suelo, en la cual según Garwood y col. (6) existe una correlación significativa entre el contenido hídrico del suelo y la eficiencia en el uso del nitrógeno, así una deficiencia de agua produce un estrés fisiológico en la planta, que disminuye la tasa de crecimiento debido a una reducción en la tasa de incremento de área foliar. Número de hojas vivas En el Gráfico 2 se presenta la evolución del numero de hojas vivas por estolón (NHV/E) a través del tiempo, observándose la diferencia entre los niveles de nitrógeno; estos resultados muestran que la aplicación de nitrógeno y el factor tiempo afectan significativamente (P<0,001) el NHV/E, pero la interacción entre estos dos factores resultó no significativa. Se realizó la prueba de medias, resultando para el caso del nitrógeno los grupos homogéneos A y B; el primero formado por el nivel N, con promedio de 32,62 hojas vivas/estolón y el segundo formado por los niveles N, y No con 23,69 y 21,17 hojas vivas/estolón, respectivamente; lo cual significa que existen diferencias significativas entre la dosis alta con respecto a las otras dos, pero no entre N, y No. En el caso del factor tiempo, presentado en el Cuadro 3 se observan tres grupos, significando que hay un incremento rápido del número de hojas vivas por la alta tasa de aparición foliar durante la primera semana (Gráfico l), que luego a partir de la segunda disminuye y en la tercera se estabiliza y permanece sin incremento significativo a partir de la cuarta semana por el fenómeno de senescencia, es decir el numero de hojas que nacen es similar al que mueren. (ver Gráfico 3 y Cuadro 4)
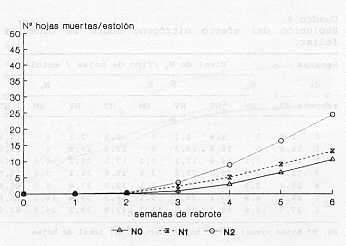
Senescencia foliar El Gráfico 3 y el Cuadro 4 muestran que a partir de la tercera semana el fenómeno de senescencia alcanza valores importantes, pero mas acentuado en el nivel No por el hecho que en este tratamiento ha habido una mayor aparición de hojas y el número de hojas que alcanza la senescencia es también mayor. Según resultados presentados por Huguet (8), en el pasto pangola (Digitaria decumbens Stent) la senescencia foliar es importante a partir del día 19 de rebrote y mas acentuado en dosis alta de nitrógeno (135 kg/ha).
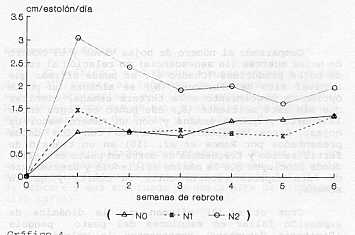
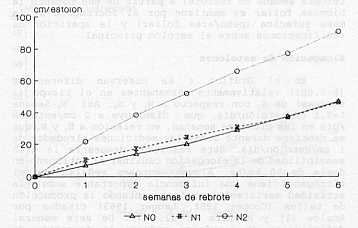
Comparando el número de hojas vivas y el número de hojas muertas (la senescencia) en relación al total de hojas producidas (Cuadro 4), se puede afirmar que a nivel alto de nitrógeno (N2) se alcanza un punto óptimo de crecimiento a la tercera semana, mientras que sin este nutriente (No) ese punto se logra entre la cuarta y quinta semana y con un número menor de hojas. Estos resultados coinciden con los datos presentados por Ramos et al. (10) en un trabajo de fertilización y frecuencia de corte en pasto estrella, donde concluyen que la máxima eficiencia y crecimiento de esta especie estaría entre la cuarta y quinta semana. Cruz et al. (3) afirman que la dinámica de expansión foliar en estolones del pasto pangola (Digitaria decumbens) representan la velocidad de elongación neta (elongación - senescencia) en las hojas principales, la cual es negativa luego de la tercera semana de rebrote; a partir de ese momento la biomasa foliar se mantiene por el incremento de la masa sufacica (pesolarea foliar) y la aparición de ramificaciones sobre el estolón principal. Elongación de estolones En el Gráfico 4 se observan diferencias (p<0,001) relativamente permanentes en el tiempo (3 semanas) de N2, con respecto a N, y No. Así N2-Semana 1=3,1 cm/estolón/día, que disminuye a 2 cm/estolón /día en las próximas semanas, en relación a N, y No que se mantiene durante todas, las mediciones alrededor de 1 cm/estolón/día. Este resultado demuestra la gran sensibilidad de la elongación caulinar al nitrógeno en dosis de 50 kg/ha. Algunos autores señalan que el nitrógeno tiene una influencia importante sobre la actividad meristernática, incrementando la producción de tallos (Cooper 1951, Langer 1963) citados por Anslow (1) y Herrera et al. (7) . De esta manera, deficiencias de nitrógeno reducen la formación de nuevos tallos.
En el Gráfico 5 se presentan los valores acumulados de la elongación de estolones, observando como se incrementan las diferencias (P< 0,001) entre N2 con respecto a N, y No en la medida como avanza el crecimiento y edad de la pastura, llegando a duplicar el valor. García (5) reportó valores de 0,57; 1,0 y 0,96
cm/estolón/día en tres semanas de observación. Resultados similares presentó Condoví
en Cuba (2), quien trabajando con varias especies de Cynodon, obtuvo una tasa de
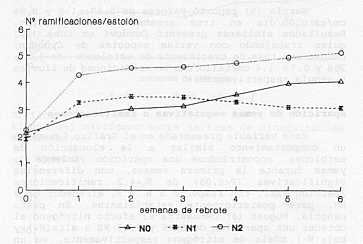
crecimiento de estolones de Aparición de yemas vegetativas o ramificaciones Esta variable presentada en el Gráfico 6 muestra un comportamiento similar a la elongación de estolones, encontrándose una aparición violenta de yemas durante la primera semana, con diferencias significativas (P<0,05) de N,=4,2 ramificaciones/estolón (RE) , con respecto a N, y N, alrededor de 3 RE, para posteriormente estabilizarse. En pasto pangola, Huguet (8) demuestra el efecto nitrógeno al obtener una aparición de 8,46 y 3,03 RE a alta (N+) y baja (N-) dosis de nitrógeno respectivamente, en un rebrote de 35 días y observa un mayor desarrollo de las ramificaciones en N+, particularmente debido a la elongación del tallo y la aparición de hojas. En síntesis, se observan diferencias altamente significativas (P<0,01) de N, en relación a N, y No sobre la tasa de aparición y dinámica foliar (número de hojas vivas), la tasa de elongación de tallos (cm/estolón/día) y la aparición de ramificaciones. Estas diferencias mantienen ese nivel de significancia solo durante las primeras 2-3 semanas; tiempo en el cual las plantas utilizan al máximo y/o permanece el efecto del nitrógeno. En cuanto a la senescencia, esta es importante a partir de la 3-4 semana, siendo mayor para el caso de N2 motivado al mayor número de hojas producidas y la mayor rapidez y/o dinámica de flujo de biomasa generada.
El conjunto de estos resultados guardan similitud a los mostrados por Cruz et al. (3) en pangola, encontrando una fuerte acumulación de la MS en el compartimiento de hojas, con un tope a la tercera semana del rebrote. El fuerte crecimiento de los estolones se inicia mas tardíamente y finalmente acumulan mayor cantidad de MS que las hojas, resaltando así el efecto del nitrógeno sobre los estolones en mayor cuantía que sobre las hojas. Este autor muestra que la senescencia foliar es mayor que el crecimiento a partir de la tercera semana. Resultados similares fueron reportados por Schemoul (13), trabajando sobre Díchantíum aristatum. CONCLUSIONES
RESUMEN La productividad de las especies forrajeras tropicales sometidas al efecto de la fertilización, normalmente se determina mediante corte y pesada, lo cual ofrece resultados globales, no permitiendo explicar la evolución y dinámica del crecimiento de tallos y hojas a través del tiempo. En el presente trabajo se analizó la respuesta de los diferentes componentes morfológicos del pasto estrella a la aplicación de tres niveles de nitrógeno (N,:O, N,:25 y N,. 50 kg de nitrógeno/ha) en un diseño de bloques al azar con dos repeticiones. Mediante identificación individual de plantas se tomaron medidas semanales (7 semanas) de elongación de estolones y utilizando marcaje con pinturas se realizaron seguimientos detallados sobre aparición, duración de vida y senescencia de hojas. Los resultados muestran diferencias altamente significativas (P<0,001) entre los promedios de tasa de aparición foliar (N2;N,=No), diferencia importante solo durante la primera semana, N,=2,25; N,=1, 4; N,=I, 2 hojas/estolón/dia (HED) ; después estas valores oscilan entre N2=1,25 y N, No=0,75 HEDO. En la dinámica foliar, expresada por el número de hojas vivas por estolón (HVE) se observan diferencias altamente significativas (P<0,01) entre N2=32,62 HVE, con respecto a N,=23,69 HVE y N,=21,17 HVE. Esta variable es afectada igualmente por el factor tiempo (semanas) , existiendo diferencias altamente significativas en la primera fase de crecimiento: semana 1=14,70 HVE, semana 2=18,97 HVE; en relación a la segunda fase de crecimiento: con valores de 26,47 hasta 32,48 HVE para la semana 3 y semana 6, respectivamente. La senescencia foliar resultó significativa a partir de la semana 3 pero mas acentuada en N2. La elongación de tallos mostró igualmente diferencias altamente significativas (P<0,01) de N2 en la semana 1=3,1 cm/estolón/día, que disminuye a 2 cm/estol6n/día en las siguientes semanas, en relación a N, y No que mantienen sus mediciones alrededor de 1 cm/estolón /día. Se concluye que el mayor efecto sobre las variables estudiadas se obtuvo con 50 unidades de nitrógeno/ha (N,), la producción foliar se estabiliza con promedio de 30 hojas a partir de las semanas 3 y 4, por el incremento de la senescencia; así mismo se produce la mayor tasa de elongación de tallos. En general se observa un efecto violento del nitrógeno entre la primera y tercera semana de crecimiento. NITROGEN FERTILIZATION EFFECTS ON THE MORPOLOGICALS COMPONENTS OF STAR GRASS (Cynodon plectostachyus) IN THE ZONE OF BAJO TOCUYO, FALCÓN STATE, VENEZUELA SUMMARY The tropical forage species productivity under the effect of fertilization normally can be determined by cutting and wheighting which offers global results and daes not allow to explain the evolution and growth dynamics through time. of stems and leaves. In the present study the reponse of the differents morphological componente of star grass under three different levels of nitrogen (N,.O, N,:25 y N,: 50 kg of nitrogen/ha) in a randomized blocks with two trails desing where used. Weakly measurements of individual plants where taken, during seven weeks the stolons elongation was measured and by marking with pain the leaf appearance, duration of life and leaf senescence where measured. The results showed very highly significant differences (P<0.001) between the means rate of leaf appearance (NoN,=N,,) during the first week, N,=2.25; N,=1.4; N,=1.2 leaves/stolon/day, after that the values oscilated between N,=1.25 and N,=0.75 leaves/stolon/day. The leaf dinamics expresed by the number green leaves per stolon showed highly significant differences (P<0.01) between N,=32.62 leaves/stolon and N,=23.69, N,=21.17 leaves/stolon. This variable is afected by time (weeks) existing highly significant differences during first phase of growth: week one=14 70 leaves/stolon, week two=18.97 leaves/stolon and 'second phase of growth: week three=26.47 leaves/stolon, week six=32.48 leaves /stolon. Leaf senescence was significativa from the week three but more important in N2- Stolon elongation showed highly significant differences (P<0.01) of N,- week one=3.1 cm/stolon/day, which Recreases to 2 cm/stolon/day in other weeks and N,, N,=1 cm/stolon/day. In conclusion, the dose N2(50 kg/ha) showed the best efféct on all variables, the means leaf production was 30 green leaves trom the week three. The be'st nitrogen effect on dinamics of growth was produced between first and third week. BIBLIOGRAFÍA 1. ANSLOW R.C. The rate of appearance of leaves on tillers of the gramineae. Herbage Abstracts. Vol 36 No. 3. pp 150-155. 1966. 2. CONDOVI E. La velocidad de crecimiento floración de algunas gramíneas tropicales. Ciencia y técnica en agricultura, pastos , forrajes. Sub-estación de pastos Granma. Habana Cuba. 5 (1):101-104. 1982. 3. CRUZ P., G. ALEXANDRE y H. BAUDOT. Cinétique de la croissance foliaire et stolonifere d, un peuplement de Dígitaria decumbens au cours de la repousse. Procc. XVI Congres International des Herbages, Nice France. pp 499-500. 1989. 4. DAVIES A. Tissue turnover in the sward. En: J. HODGSON, R. D BAKER, A. DAVIES, A. S. LAIDLAN y J.D. LEAVER (Eds.) Sward Measurement handbook, Berkshire. British Grassland Society. pp 179- 208. 1981. 5. GARCIA A. Contribución al Estudio de la Dinámica del Crecimiento del Pasto Estrella (Cynodon plectostachyus, PILGER) en la Región Bajo Tocuyo, Estado Falcón. UNEFM. 54 p. 1990. (Tesis de Grado). 6. GARWOOD E., J. SALETTE y G. LEMAIRE. The influence of water sypply to grass on the response to fertilizar nitrogen and nitrogen recovery. Procc. Int. Symp. Eur. Grassland. ed. on the role of nitrogen in intensiva grassland production. wageningen. pp 59-65. 1980. 7. HERRERA R., S., N. RAMOS y Y. HERNÁNDEZ. Respuesta de la Bermuda Cruzada a la Fertilización Nitrogenada y Edad del Rebrote. V. Rendimiento de Materia Seca, Hojas, Proteína Cruda y Eficiencia de Utilización del Nitrógeno. Rev. Cubana Ciencias Agric., 20:193-201. 1986. 8. HUGUET J.M. Contribution a l'etude de la croissance d' une graminée fourragere tropicale stolonifere perenne: le pangola (Dígítaria decumbens Stent). Memoire Diplome d, Agronomie Approfondie. ENSAINRA. France. 56 p. 1989. 9. MARTINEZ D. Diagnóstico Agroecológico Bajo Tocuyo, Identificación, Verificación y Ajuste de Unidades Agroecológicas. Fonaiap-Falcón. 39 pp. 1983. (Mimeo). 10. RAMOS N., P. CURBERO y R.S. HERRERA. Edad de Rebrote y Niveles de Nitr6geno en Pasto Estrella (Cynodon nlemfuensis) . Rev. Cubana Ciencias. Agrícolas, 14:83-93. 1980. 11. ROMERO C. Contribution a I'étude de la croissance de la fetuque elevée (Festuca arundinacea SCHREB) sous páturage continu par des moutons. Approche morphogénétique. Univ. Paris XI. Orsay. France. 33 pp. + annexes. 1987. (Tesis Maestría). 12. ROMERO C., A. GARCÍA y R. FLORES. Efecto de la Fertilización Nitrogenada sobre los Componentes morfológicos del Pasto Estrella. En: Resúmenes Coloquio Internacional sobre el Estudio de la Dinámica Productiva en Pastos Estoloníferos Tropicales. Ed. Carlos Romero. Fonaiap-Falcón. Coro. Venezuela. pp 16-17. 1992. 13. SCHEMOUL E. Productivité primaire et morphogénese d' un peuplement de Petit Foin (Dichantium sp Will) en Guadeloupe. memoire Diplome d,Agronomie Approfondie. ENSA-INRA. France. 56 pp. 1988. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||