|
Zootecnia Tropical 22(2):147-156. 2004 Acumulación y distribución de biomasa en Leucaena leucocephala (lam) de Wit., durante la fase de establecimiento. I. Repartición de biomasa Eunice Guevara1 y Orlando Guenni2 [1]Instituto
Nacional de Investigaciones Agrícolas (INIA) Centro de
investigaciones agropecuarias. Recibido
y
aceptado dentro del contexto del XII Congreso de Producción
e Industria Animal 2004. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RESUMEN En el presente estudio se comparó el patrón de acumulación de biomasa aérea y radical durante los primeros 110 días después de la germinación (DDG), en tres líneas de Leucaena leucocephala (CIAT 7984, 9438, 18477). El cv. Perú se utilizó como testigo con fines de comparación. El experimento se llevó a cabo en condiciones de invernadero, cultivándose las plantas en tubos de PVC de 1 m de alto por 0.20 m de ancho. El diseño del experimento fue un completamente aleatorizado, en un arreglo factorial de 4 líneas x 5 cosechas (15, 30, 45, 60 y 110 DDG) x 5 repeticiones. Los datos fueron analizados por vía no paramétrica. Para estimar el patrón de acumulación y distribución de biomasa se midió en cada cosecha el peso seco acumulado de hojas (MSh), tallos (MSt) y raíces (MSr). A los 110 DDG, todas las líneas en general destinaron una mayor proporción de asimilados en producir hojas a expensa de tallos y raíces. La MSh no fue diferente (P<0,05) entre las líneas para las dos ultimas cosechas (60 y 110 DDG), la Mst de la línea 9438 y el cv Perú fueron mayores (P<0,05) en comparación a la línea 18477, y la MSr de esta última (18477) no fue diferente estadísticamente (P<0,05) a los 30 y 110 DDG, lo que sugiere un potencial en condiciones limitadas. La relativa mayor acumulación de biomasa en la línea 18477, justificaría su inclusión en futuros programas de selección y mejoramiento agronómico. Palabras clave: Acumulación de materia seca, distribución de biomasa, Leucaena leucocephala. Accumulation
and distribution of biomass in Leucaena
leucocephala (Lam.) de wit during the phase of establishment. II.
Repartitioning of biomass SUMMARY In the present study, dry matter accumulation and the pattern of distribution between aerial and bellow-ground biomass were studied in three accessions (CIAT 7984, 9438, 18477) of Leucaena leucocephala during 110 days after germination (DAG). Cv. Perú was used as a control plant. The experiment was carried out under glasshouse conditions, where plants were grown in PVC pots (1 m height x 0.20 m diameter). A completed randomized designed was used, in a factorial arrangement of 4 plant materials x 5 harvests (ie. 15, 30, 45, 60 and 110 DAG) x 5 replicates. To estimate biomass accumulation and assimilates distribution within the plant, harvests of individual plants were performed and the dry weight of leaf (LDM), stems (SDM) and root (DMR) at different depths was recorded. Initially, all accessions diverted a high proportion of the available energy in producing root biomass, while by the end of the establishment period relatively more photosynthetic material was used to increase leaf and stem biomass. The MSh was not different (P<0.05) among the lines for two finish court (60 and 110 DDG), the Mst of the line 9438 and the cv Peru was bigger (P<0.05) in comparison to the line 18477, and the MSr of this last one (18477) it was not different statistically (P<0.05) to the 30 and 110 DDG, what suggests a potential under non limited conditions. The distinctive pattern of distribution of forage biomass within the accession 18477, disserves further research in future programs of agronomic improvement. Key words: Dry matter accumulation, biomass distribution, Leucaena leucocephala. INTRODUCCIÓN Leucaena leucocephala es utilizada estratégicamente como recurso alimenticio en unidades de producción animal, por su multiplicidad de uso como árbol forrajero y madera. Entre sus bondades se destacan: altos contenido de proteína cruda, vitaminas, minerales, carotenos y fibra que determinan su alto valor nutritivo; por otro parte tolera periodos moderados de sequía, y tiene capacidad de adaptarse a suelos medianamente ácidos (pH 5.0 a 6.4; Shelton H. y R. Jones. 1994; Cáceres y Santana, 1991). Esta especie de alto potencial productivo es considerada una alternativa para mejorar las explotaciones pecuarias en el trópico (Clavero, 1996). Los distintos valores encontrados en la literatura en cuanto a producción de materia seca, relación hoja tallo, valor nutritivo y grado de adaptabilidad a distintos ecosistemas, entre y dentro de especies del género Leucaena spp. (Camacaro et al, 2002; Guevara, 1998; Davila et al, 1996; Razz R., T. Clavero y J Pérez. 1994), podrían estar íntimamente relacionados a una alta variabilidad interespecífica e intraespecífica de la especie. Diferencias en el patrón de acumulación de biomasa y repartición de asimilados entre el vástago y la raíz podrían igualmente determinar un mayor o menor potencial adaptativo a determinadas condiciones edafoclimáticas y de manejo. Por consiguiente, es de un gran interés biológico y agronómico conocer el potencial genético de variación dentro de la especie en relación al crecimiento y distribución de biomasa. El objetivo del presente trabajo fue comparar el patrón de acumulación y distribución de biomasa aérea y radical entre cuatro líneas contrastantes en rendimiento de materia seca de L. leucocephala durante el período de establecimiento. MATERIALES Y MÉTODOS Ubicación
y descripción del sitio experimental El experimento se realizó en el invernadero del Centro Nacional de Investigaciones Agropecuarias (CENIAP), ubicado en Maracay, estado Aragua (10° 17’ LN y 67° 37’ LO). La temperatura media máxima fue de 39 ºC y la mínima de 19 ºC. Material
vegetal y condiciones de cultivo. Variables medidas. Se seleccionaron tres líneas de Leucaena: CIAT: 7984, 9438 y 18477, provenientes de Colombia, más el cv Perú como testigo. Detalles acerca de los criterios de selección de estas líneas son dados por Guevara (1998). Su escogencia se basó en la respuesta diferencial que las mismas han mostrado en relación a la producción de materia seca en condiciones de campo. Las plantas de cada línea fueron cultivadas en tubos de PVC de 1 m de alto x 0.20 de diámetro. El sustrato utilizado y las condiciones de crecimiento y cosecha fueron descritos por Guevara (1998). La variables biométricas medidas en cada cosecha o muestreo fueron: 1) Materia seca (g) en el vástago (MSv), obtenida a partir de la sumatoria de materia seca de las hojas (MSh) y del tallo (MSt) y 2) materia seca radical (MSr). Esta última representó la sumatoria de la biomasa de raíces obtenidas de los estratos: 0-5, 5-25, 25-50 y 50-75 cm de profundidad. La MS total (g) de la planta fue la sumatoria de Msv más MSr. La relación raíz:vástago (R:V): se obtuvo a partir del cociente MSr/MSv. Diseño
del experimento y análisis estadístico de los resultados. Se utilizó un diseño completamente al azar, en un arreglo factorial de 4 líneas x 5 cosechas (15, 30, 45, 60 y 110 DDG) x 5 repeticiones. Los datos fueron analizados por vía no paramétrica con el análisis de varianza de Kruskal y Wallis, cuya distribución es Chi-cuadrado (X2; α=0.05). Para las pruebas de medias se utilizó la diferencia de medias de los rangos de cada tratamiento entre la desviación estándar del error (Q 0.05,k-1; Jerrold, 1984). RESULTADOS Y DISCUSIÓN Patrón
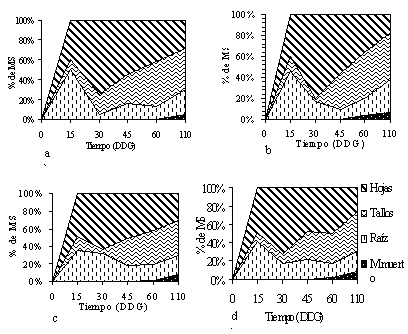
de distribución de biomasa. El patrón de distribución de biomasa refleja el gasto energético invertido por la planta para la producción de los distintos órganos de la planta (Shiabata et al, 1995). En la Figura 1 se muestra para cada línea, la distribución relativa de la MS (gg-1 de peso seco total x 100) de los distintos órganos de la planta y la materia muerta (senescente) a lo largo del tiempo. En general, todos los materiales evaluados (CIAT: 7984, 9438 y 18477) incluyendo el cv Perú, presentaron un comportamiento similar (P>0.05) en la distribución de biomasa, destinando en los primeros 15 días hasta un 40% de la energía en la producción de raíces. Esta inversión decae a un 10-30% a los 30 DDG, debido a la rápida acumulación de superficie asimilatoria. La alta inversión inicial de energía en MSr observada en la especie, es importante para garantizar el éxito durante el establecimiento, sobre todo si hay limitaciones edáficas (de pH, altos contenido de aluminio, deficiencia de Fósforo, entre otros) y/o competencia por espacio, luz y agua (Jaafar et al 1993). A los 30 DDG, la proporción de hojas es máxima en todas las líneas, alcanzando un 60-80% de toda la planta. A partir de los 45 DDG, todos las líneas incluyendo el cv Perú ven disminuidas (P>0.05) sus inversiones energéticos en la producción de raíces y, en menor grado, de hojas; esto debido principalmente a la alta competencia de los órganos de soporte aéreo (tallos y ramas). Entre los 45 y 110 DDG, la proporción de tallo y ramas osciló entre un 25 y 35% en todas las líneas evaluadas, mientras que para el sistema radical dicho rango fue 20-25%.
El material senescente (ramas y hojas secas) comenzó a desprenderse de la planta a partir de los 60 DDG, alcanzando para el final del experimento un valor de alrededor de un 10% en todas las líneas. Se observó floración solo en las línea 17477 y en el cv Perú, correspondiendo la biomasa de estructuras reproductivas a un 2 % del total de la planta. A los 60 DDG, las líneas consideradas de alto rendimiento (CIAT: 7984 y 9438) presentaron un 40 % de tallos, mientras que la línea 18477 y el cv Perú alcanzaron sólo un 30 %. Aún cuando estas diferencias en los valores porcentuales pudieran ser significativas, los valores absolutos de MS (g planta-1) de hojas y tallos para esa fecha no presentaron diferencias significativas. A los 110 DDG, las líneas 7984 y el 18477 destinaron mayor energía (35-40% vs. 20-25%) en la producción de hojas. Aún cuando las líneas 18477 y el cv Perú mostraron una relativa menor producción de raíces laterales, esto se vió aparentemente compensado por un mayor crecimiento (g planta-1) de la raíz principal. Patrón
de acumulación de biomasa. A
todo lo largo del experimento, el peso seco total de planta no mostró
diferencias significativas (P>0.05) entre los materiales estudiados.
Sin embargo, la acumulación de MS en los distintos componentes de la
planta si mostró diferencias significativas entre líneas, las cuales,
fueron dependientes del estado de crecimiento (Cuadro 1). En este
sentido, la biomasa foliar (MSh) por ejemplo mostró diferencias
significativas (P<0.05) a los30 y 45 DDG, siendo mayor en el cv Perú.
A los 110 DDG, estas diferencias se hacen mínimas y no significativas
(P:0.05), teniendo la MSh un rango de variación de 17 a 23 g planta-1.
La MSt fue solo significativamente
diferente (P<0.01) entre los materiales sólo a los 110 DDG, siendo
mayor en el cv Perú y la línea 9438 (37-48 g planta-1) que
en las otras líneas (24-28 g planta-1) (Cuadro
1). Como consecuencia, la relación hoja:tallo (MSh:MSt) fue
significativamente mayor (P<0.05) en la línea 18477 (0.72) que en la
línea 9438 (0.35) (Cuadro 1).
Un aumento relativo del peso de tallos en la planta debido a la lignificación de los mismos, indica la existencia de una trasformación fisiológica, de un estado joven a maduro (Humphreys, 1991). El poseer una menor proporción de estructuras lignificadas pudiera representar una ventaja desde el punto de vista agronómico, dado que este tipo de material biológico con una mejor calidad forrajera, como fue el caso de la línea 18477, se usaría mayormente en la alimentación de becerros y de pequeños herbívoros (Siregar, 1982), considerando desde luego la tolerancia de estos animales a la Mimosina, ß-N [3-(hydroy-4 pyridone)]-oc-aminopropionic acid, estimaciones de consumos entre 0.2 a 0.3g de mimosina por kg de peso vivo en ovejos, mostraron efectos tóxicos (Ravindran, 1979) y 2 a 3 % de peso vivo para becerros en amamantamiento restringido y a pastoreo (Baez, et al.,1983). La biomasa total de raíces (MSr) fue similar entre las líneas hasta los 30 DDG (Cuadro 1). Hasta los 60 DDG, el rango de variación en el peso seco de la raíz fue muy estrecho (4-6 g planta-1), representado alrededor de un 15% del peso total de la planta. Es solo a los 110 DDG que aumenta la proporción de la raíz en la planta y las diferencias entre especies se hacen mas marcadas la los 110 DDG. En este caso, la línea 18477 mostró una menor biomasa radical (13.1 g planta-1) en comparación con la línea 9438 (22.9 g planta-1) (Cuadro 1). Un baja biomasa radical podría limitar el acceso al agua y nutrientes en el suelo, lo cual podría a su vez limitar el crecimiento del vástago. Sin embargo, fue este línea la única en florecer (a los 70 DDG), lo cual pudo haber inducido a una disminución en la acumulación de biomasa en los restantes órganos de la planta. La disminución en el peso radical estuvo asociada a un menor desarrollo de raíces laterales. Las diferencias observadas entre líneas en relación a la biomasa radical se evidenciaron también a nivel de la distribución del peso de la raíz a lo largo del perfil de suelo (Cuadro 2). Aún cuando la línea 18477 y el cv. Perú fueron las primeras en producir raíces entre 25 y 50 cm, al final del experimento fueron las líneas 7984 y 9438 las que desarrollaron una mayor biomasa de raíces entre 50 y 75 cm (2.6-2.8 g planta-1 vs. 1.1-1.8 g planta-1) (Cuadro 2). En todas las líneas mas del 50% de la biomas radical se distribuyó entre los primeros 25 cm, oscilando este valor entre 66% (línea 9438) y 83% (línea 18477 y cv. Perú). Esto implica que, al menos que ocurra un cambio marcado en la distribución radical debido al stress hídrico, el resto de la biomasa de raíces debe suplir las necesidades de agua y nutrimentos a medida que ocurre el secamiento del suelo durante la sequía (Roundy et al 1993). A partir de los 110 DDG, la raíz crece acumulando mayor densidad de longitud radical entre los 50 a 75 cm de profundidad, observándose engrosamiento de la raíz principal y las RL. Para las líneas 7984 y 9438, el poseer un relativo mayor desarrollo radical con la profundidad, pudiera representar entonces un mecanismo efectivo para evadir el déficit hídrico y así mantener una mayor producción de biomasa en el año (Russell, 1992). Los resultados obtenidos confirman que algunos materiales de L. leucocephala se caracterizan por una alta producción de biomasa aérea, teniendo sus raíces igualmente una buena capacidad de crecer rápidamente garantizando así un rápido establecimiento, lo que se refleja probablemente en una mayor eficiencia de utilización de los recursos (agua y nutrimentos) para la producción de órganos fotosíntetizadores. (Jaafar et al., 1993; Ansari et al,1995). Relación
raíz: vástago (R:V) Este cociente disminuyó progresivamente (P>0.05) hasta los 60 DDG en todas las líneas, variando desde 0.73-1.04 (15 DDG) hasta 0.14-0.21 (60 DDG). Para el final del experimento (110 DDG) dicho cociente aumentó ligeramente en todas las líneas, con un rango de variación de 0.32-0.42. Este aumento en la relación R:V pudo haber ocurrido debido a la ocurrencia simultánea de dos eventos en la planta: 1) Una disminución relativa en la proporción de hojas y 2) un aumento relativo en la fracción total de raíces. Valores en general mas altos en el cociente R:V fueron encontrados por Espinoza et al. (1997), quienes observaron utilizando distintos niveles de roca fosfórica, un rango de 0.77-1.25 de relación entre la parte área y radical (V:R) raíz vástago para la línea 9438. Sin embargo, a diferencia de este estudio, dicho cociente no mostró una disminución en el tiempo. CONCLUSIONES Todos los materiales estudiados, incluyendo el cv Perú, presentaron un comportamiento similar en la distribución de biomasa antes de los 60 DDG, destinando gran cantidad de energía en la producción de raíces. Este patrón de distribución de asimilados que favorece a la raíz, es importante para garantizar el éxito durante el establecimiento, sobre todo si hay limitaciones edáficas y/o competencia por nutrimentos y agua. Por otra parte, al final del establecimiento la especie destina mayor cantidad de recurso fotosintético en producción de hojas de hojas que tallos y raíces, lo que le proporciona en consecuencia un incremento sostenido en la producción de biomasa forrajera La línea 18477, mostró un comportamiento similar a las líneas consideradas de alto rendimiento en campo, pero con una relativa mayor proporción de materia seca de alto valor nutritivo. Esto apunta a la necesidad de reconsiderar los resultados de evaluaciones de campo donde se comparan distintos materiales, de manera que se incluyen las distintas fracciones de la biomasa potencialmente forrajera. En términos de producción de materia seca, la accesión 18477 parece poseer un alto potencial genético, lo que justificaría su inclusión en futuros programas de selección y mejoramiento agronómico. BIBLIOGRAFÍA Ansari
S., Kumar P. y B. Gupta. 1995.
Root surface area based on absorption and desorption of nitrite. Plant
and Soil 175: 133 - 137. Argenti P. y F. Espinoza. 1993. Leucaena. Leucaena leucocephala. Serie B. FONAIAP, CENIAP. Maracay: 19. Baez L.; B. Pound y G. Peña. 1983. Nota sobre el comportamiento de becerros alimentados con Leucaena leucocephala (Lam.) de Wit. Producción Animal tropical, 8:64-66. Caceres O. y H. Santana. 1990. Valor nutritivo de Leucaena leucocephala cv cunningam en diferentes momentos del Año. Rev. Pastos y Forrajes 13:197-201. Clavero, T. 1996. Las leguminosas forrajeras arbóreas: Sus perspectiva para el trópico americano. In Clavero, T. (Ed) Leguminosas Forrajeras Arbóreas en la Agricultura Tropical. Centro de Transferencia de Tecnología en pastos y Forrajes Universidad del Zulia. 1-10 pp. Dávila C. y D. Urbano. 1996. Evaluación de ecotipos de Leucaena (Leucaena leucocephala) bajo corte en el Sur del Lago de Maracaibo. Revista Facultad de Agronomía (LUZ), 13(5):539-550. Espinoza F., Díaz Y., and P. Argenti. 1997. The effect of rockphosphate on Leucaena leucocephala growth in acids soils of Venezuela. Leucnet New . 4:24-26. Guevara E. 1998. Acumulación de biomasa aérea y patrón de crecimiento radical en cuatro ecotipos de Leucaena leucocephala Lam de Wit., durante la fase de establecimiento bajo condiciones de invernadero. Tesis de M. Sc. UCV-Fac. Agronomías y Fac. de Ciencias Veterinaria, Maracay, Venezuela. 124 p. Humphreys L. R. 1991.
Effects of defoliation on the growth of tropical pastures. Tropical
Pasture Utilitation. Cambridge University Press. pp. 46-65. Jaafar
M. N., Stone L.R. y D. E Goodrum. 1993. Rooting depth and dry matter
development of sunflower. Agron. J., 85: 281 - 286. Jerrold
H. 1984. Biostatistical Analysis. Second Edition. Prentie-Hall
International. Inc. Northern Illinois University: 718 p. Ravindran, V. 1979. Leucaena leucocephala (Lam.) de Wit a poder legume with a future. Rev. Tropical Agriculturist, vol 85:70-82 Razz R., T. Clavero y J Pérez. 1994. Crecimiento y rendimiento de materia seca de dos ecotipos de Leucaena leucocephala bajo diferentes niveles de fertilización. Revista Facultad de Agronomía (LUZ), 11(4):347-357. Roundy B. A., Winkel V. K., Cox J. R., Dobrenz A. K. y Tewolde H., 1993. Sowing depth and soil water effects on seedling emergence and root morphology of three warm-season grasses. Agronomy Journal. 85 (5):975-982. Russell W. 1992. Crecimiento y Funcionamiento de las raíces. En: Condiciones del suelo y desarrollo de las plantas. Ed Mundi, Prensa Madrid: 1045 p. Shelton
H. y R. Jones. 1994.
Opportunities and limitations in Leucaena.
Shelton, H.; Piggin, C. y Brewbaker, J. (Eds) Leucaena.
Opportunities and limitations. Proceeding. Bogor, Indonesia. 16-22 pp. Shiabata
O., Kinoshita T. y T. Arai.
1995. Dry mass
production of several plants grown in habits differing in altitude. J
Fac. Sci., 10:41-46. Siregar A. P. 1982. The prospect of “Lamtoro” (Leucaena) as feed for livestock and poultry production in Indonesia. Ind. Agric. Res. Dev., 4(4):98-1 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||