|
Zootecnia Tropical 23(4):411-427. 2005 Comportamiento reproductivo de vacas Brahman de primera lactancia suplementadas con proteína no degradable Belkys J. Vásquez M.1* y Pedro Bastidas M.2
1
Instituto Nacional de
Investigaciones Agrícolas. 2
Universidad Central de Venezuela. Recibido: 16/08/05 Aceptado: 21/11/05 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Resumen Para evaluar el efecto del consumo de dos niveles de proteína no degradable (harina de pescado) en el rumen (PNDR) sobre el comportamiento reproductivo postparto, 30 vacas Brahman de primera lactancia fueron asignadas al azar a tres grupos. El grupo A (n=10) recibió 40% de PNDR, el grupo B (n=10) recibió 60% de PNDR y el grupo C (n=10) no recibió suplementación. La suplementación comenzó 60 días antes del inicio de la temporada de servicios (TS). Los animales se mantuvieron en potreros con pasturas de Panicum maximum, Brachiaria mutica y Cynodon plectostachyus. Las vacas y los becerros fueron evaluados quincenalmente para determinar ganancia diaria promedio (GDP) y condición corporal (CC). Dos veces por semana se realizaron ecosonografías transrectales para la evaluación los folículos presentes, además de extraerse 10 ml de sangre para la cuantificación de progesterona. Los grupos A y B presentaron una mayor GDP al inicio de la TS (P<0,04). El grupo A tendió (P<0,1) a una mejor condición corporal al inicio de la TS comparado con el grupo control. A los 30 y 60 días de temporada, el grupo A se encontraba en mejor condición corporal (P<0,02) y (P<0,08), respectivamente, comparado con el grupo control. El grupo B tendió (P<0,1) a presentar una mayor proporción de celos durante el primer mes de TS, comparado con el grupo A y Control. En el segundo mes de TS, el número de celos fue mayor (P<0,02) y el intervalo inicio de suplementación- primer celo fue menor (P<0,02) para el grupo B comparado con el grupo Control. El número de folículos clase 1 y clase 2 no fue afectado por los tratamientos (P>0,05). El número de folículos clase 3 fue mayor (P<0,04) en el grupo B, comparado con el grupo A y Control. La suplementación estratégica con PNDR, previo al inicio de la temporada de servicios, puede mejorar el comportamiento reproductivo de vacas Brahman de primera lactación. Palabras clave: Proteína no degradable, vacas Brahman, anestro postparto, folículos ováricos. Reproductive performance of first lactating Brahman cows supplemented with undegradable protein SUMMARY To evaluate the effect of the two undegradable protein levels (UDP) in the diet on reproductive performance, 30 first-calf Brahman heifers were assigned at random to three groups. Group A (n =10) received 40% of UDP, group B (n= 10), received 60% of UDP, and group C (n=10) without any supplementation. The supplementation started 60 d before the beginning of the breeding season (BS). The animals were maintained on pastures of Panicum maximum, Brachiaria mutica and Cynodon plectostachyus. Cows and calves were evaluated biweekly to determine average daily gain (ADG) and body condition score (BCS). Ultrasound was performed twice a week and blood samples (10 mL) were collected to determine progesterone. Cows in groups A and B had a greater ADG at the beginning of the BS (P<0.04) compared to control cows. Also animals in group A did tend (P<0.1) to have better BCS at the beginning of the BS than cows in control group. Furthermore, 30 and 60 d from the beginning of the BS, cows from group A had better BCS (P<0.02 and P<0.08, respectively), compared to the control group. Cows of group B tended (P<0.1) to have a greater proportion of estrus during the first month of BS compared to the group A and control group. During the second month of BS, the number of cows in estrus was greater for group B (P<0.02) than in cows of the control group. The interval from supplementation to first estrous was shorter for group B (P<0.02). The number of class 1 and class 2 follicles were not affected by treatment (P>0.05). However, the number of class 3 follicles was greater (P<0.04) in group B than in A and control groups. The strategic UDP supplementation, before the beginning of the breeding season, can improve the reproductive performance of first-calf Brahman heifers. Keywords: Undegraded protein, Brahman cows, ovarian follicles, anestrous postpartum. INTRODUCCIÓN Uno de lo factores que afectan la eficiencia reproductiva del ganado Bos indicus en condiciones tropicales, lo constituye el comportamiento reproductivo de las vacas Brahman de primera lactancia, las cuales presentan intervalos entre partos muy prolongados debido a la mala nutrición y al manejo (Olivares, 1981; Plasse, 1988). Por otra parte, la implementación de un temporada de monta restringida y las deficiencias alimenticias críticas, hacen muy difícil el reinicio de la ciclicidad estral de vacas Brahman, particularmente las de primer parto. Estas condiciones de estrés son igualmente limitantes para alcanzar una alta fertilidad en el rebaño. Estudios previos han sugerido que la suplementación proteica puede mejorar la reproducción del ganado de carne (Nolan et al., 1988). Investigaciones posteriores reportaron los beneficios de la incorporación de proteína no degradable en el rumen (PNDR). En este sentido, la NRC (1996) resaltó que el cálculo de los requerimientos proteicos en el ganado de carne debe incluir los requerimientos de proteína degradable en el rumen (PDR) y los requerimientos de PNDR. Wiley et al. (1991) y Anderson et al. (2001) concluyeron que después de cubrir los requerimientos de proteína degradable en el rumen, el aporte de proteína adicional como proteína no degradable en el rumen puede disminuir la duración del anestro postparto, además de mejorar la tasa de concepción (Triplett et al., 1995). Esto puede ser explicado por el hecho de que los mediadores del balance energético y la función reproductiva son influenciados por la suplementación proteica (Wiley et al., 1991; Noguchi, 2000; Hess et al., 2005). El eje hipotálamo-hipófisis-ovario tiene un rol fundamental en la integración del estatus nutricional y la función reproductiva (Schillo, 1992; Wiltbank et al., 2002). En este sentido, Richards et al. (1989) asociaron la subnutrición con la disminución de la secreción de las hormonas FSH y LH. Estudios más recientes realizados por Kane et al. (2002) reportaron el aumento de la secreción de LH inducida por GnRH, después de la suplementación con PNDR. Por otra parte, Kane et al. (2004) sugirieron que las variaciones del comportamiento reproductivo debido al consumo de PNDR en vacas de carne, puede estar asociado con cambios en la síntesis, almacenamiento y secreción de gonadotropinas en la glándula pituitaria anterior y/o dinámica folicular. En nuestro país existe poca información con respecto a la suplementación de vacas de carne con PNDR, por consiguiente el objetivo de este estudio fue investigar el efecto de la adición de PNDR en la dieta, previo al inicio de la temporada de servicios, sobre el comportamiento productivo y reproductivo de vacas Brahman de primera lactancia. MATERIALES Y MÉTODOS El estudio se realizó en la Estación Experimental La Cumaca, perteneciente a la Facultad de Ciencias Veterinarias, Universidad Central de Venezuela, ubicada en San Felipe, estado Yaracuy. Esta Estación se ubica en una zona de bosque seco a bosque húmedo tropical con dos estaciones climáticas bien definidas, de enero a mayo la estación seca y desde la segunda mitad de mayo a septiembre, la de lluvia. La precipitación anual es de 1.868 mm, una altitud aproximada de 130 msnm y una temperatura promedio de 26-28ºC. Se utilizaron 30 vacas Brahman registradas, de primer parto, las cuales parieron entre los meses de enero a abril. Al momento del parto fueron asignadas al azar a tres grupos de suplementación. El grupo A recibió 2 kg/animal/día de un suplemento basado en harina de pescado con un 40% de PND, el grupo B recibió 2 kg/animal/día de suplemento con un 60% de PND y el grupo C no recibió suplementación (grupo control). Todos los animales recibieron mezclas minerales completas.
La base de la ración experimental fue una dieta isoenergética (73% NDT) y con un contenido de proteína cruda (PC) del treinta por ciento (30%) para la dieta A y del treinta y cinco por ciento (35%) para la dieta B (Cuadro 1). La ración base fue la misma para los dos grupos de animales y aportó harina de pescado, afrecho de trigo, melaza y harina de algodón (Cuadro 1).
La suplementación se suministró diariamente y comenzó 60 días antes del inicio de la temporada de servicios hasta 15 días después de iniciada la temporada (Figura 1). Todos los animales fueron mantenidos en pasturas tipo Panicum maximum, Brachiaria mutica y Cynodon plectostachyus, siendo separados al momento de recibir el suplemento. Estudios previos realizados a los pastizales, muestran que el contenido de PC varió de 5 a 7,7%. Las vacas y los becerros fueron evaluados cada dos semanas para determinar ganancia diaria promedio (GDP), condición corporal (CC). Para la medición de la condición corporal, se utilizó la escala del uno al nueve, donde uno es emaciado y nueve es obeso (Wagner et al., 1988). Dos veces por semana se realizaron palpaciones y ecosonografías transrectales para la evaluación del tamaño y número de los folículos presentes. Se utilizó un equipo de ultrasonido tiempo real, modelo lineal, equipado con una sonda rectal de 7,5 MHz. Los folículos fueron clasificados de acuerdo a lo establecido por Díaz et al. (1998), de la siguiente manera: clase 1 (folículos ≤ 5 mm); clase 2 (folículos de 6 a 9 mm); clase 3 (folículos ≥ 10 mm). Fue definido como folículo dominante, un folículo mayor de diez milímetros presente en el ovario, en ausencia de otros folículos grandes (Murphy et al., 1990). Además dos veces a la semana se tomaron 10 mL de sangre por venipuntura coccígea para la cuantificación de progesterona (P4). Estas muestras fueron centrifugadas a 3.000 rpm durante 10 minutos y posteriormente congeladas a -20ºC hasta su procesamiento por radioinmunoanálisis (RIA) de fase sólida sin extracción. La determinación de P4 se realizó utilizando kits suministrados por la Agencia Internacional de Energía Atómica (Viena, Austria). Se consideró actividad luteal niveles de P4 superiores a 1 ng/mL (Bishop et al., 1994) Sesenta días después de iniciada la suplementación, todos los animales ingresaron a una temporada de servicios de cinco meses (Figura 1). La detección de celo se realizó por observaciones directas diarias, dos veces al día, complementadas con toros receladores epidididectomizados y con desviación de pene, provistos de un arnés marcador. A los diecinueve días de iniciada la temporada de servicios, todos los animales fueron sometidos a un régimen de amamantamiento restringido. Un animal del estudio presentó valores de P4 mayores a 1 ng/mL al inicio del ensayo y fue excluido del experimento. Otro animal fue excluido por muerte del becerro. El diseño aplicado fue completamente aleatorizado, donde los datos obtenidos de: intervalo inicio de suplementación - primer celo (IPC), número de servicios por concepción (NSC), ganancia diaria promedio (GDP), cambios de condición corporal (CCC), número de folículos totales clase 1, clase 2 y clase 3 (NFT), fueron ajustados para grupo de estudio y se analizaron a través del método de mínimos cuadrados. En el caso de la variable número de folículos clase 3, por no seguir una distribución normal, se realizó una transformación de raíz cuadrada. Para el análisis de CCC, se utilizó la prueba de Kruskal-Wallis para datos no paramétricos. Para el análisis de IPC, NSC, GDP, CCC y NFT se incluyó en el modelo el efecto del tratamiento únicamente. El efecto del tratamiento sobre el porcentaje de celos durante el segundo mes y tercer mes de la temporada de servicios, sobre el porcentaje de preñez total, así como el efecto del tratamiento sobre el porcentaje de animales con folículo dominante, fue comparado usando pruebas de chi cuadrado. RESULTADOS Y DISCUSIÓN A los primeros treinta días, después del inicio de la suplementación, no se observó diferencias en GDP entre los grupos y éstos mantuvieron una disminución mínima de peso (Figura 2). A los sesenta días del ensayo se observa una ganancia de peso a favor de los grupos suplementados (P<0,04). Estos resultados concuerdan con los datos aportados por Anderson et al. (2001), quienes concluyen que la suplementación con PND redujo la pérdida de peso durante el periodo de lactancia en vacas de carne de primer parto. Para los meses restantes del ensayo no se observan diferencias entre los grupos. La GDP de los becerros no fue afectada por los tratamientos (P>0,05), lo cual coincide con lo expuesto por Alderton et al. (2000) y Strauch et al. (2001).
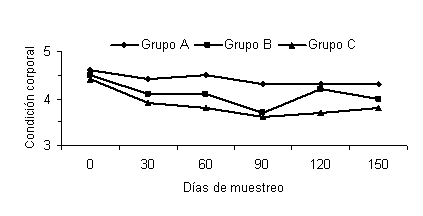
En cuanto a la condición corporal, hubo una tendencia (P<0,1) a una mayor condición corporal en los animales del grupo A, comparados con los animales del grupo control, al inicio de la temporada de servicios (TS) (Figura 3). Este grupo igualmente tuvo una mejor condición corporal a los 30 (P<0,02) y 60 días (P<0,08) de iniciada la TS, comparado con el grupo control. Estos resultados son comparables a los reportados por Alderton et al. (2000), al suministrar PNDR conjuntamente con PDR. Similarmente, Strauch et al. (2001), encontraron que a medida que avanzaba el periodo postparto, las vacas suplementadas con PNDR presentaron una mayor condición corporal comparadas con las vacas que no la recibieron. En los meses restantes, de la TS no se observaron diferencias.
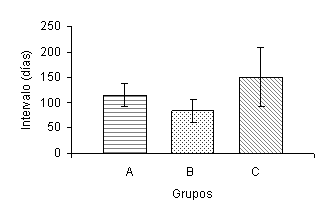
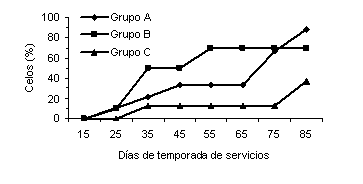
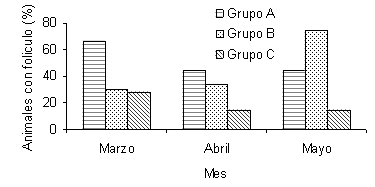
Para la variable intervalo inicio de suplementación - primer celo se observaron diferencias (P<0,02) entre los tratamientos (Figura 4). Las vacas del grupo B presentaron un intervalo al primer celo postparto más corto que las vacas del grupo A y control. Dhyuvetter et al. (1993) reportaron que los animales que recibieron un menor porcentaje de PNDR, retornaron al estro más temprano. Investigaciones posteriores han encontrado que el intervalo al primer celo no es afectado por el aporte de PNDR (Alderton et al., 2000; Strauch et al., 2001). Sin embargo, Ciccioli et al. (2003) reportaron que el intervalo al primer estro es más corto para aquellos animales que reciben una alimentación que les permite ganar más peso. A pesar de que la tasa total de estros al final de la temporada de servicios no fue afectada por los tratamientos, las vacas del grupo B presentaron una mayor proporción de estros (P<0,02), comparadas con las vacas del grupo control durante los primeros 60 días de la TS (Figura 5). La proporción acumulada de hembras en celo durante el tercer mes de TS fue mayor (P<0,05) para el grupo A, con respecto al Control (Figura 6).
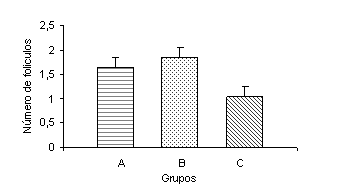
La tasa de concepción no fue afectada por los tratamientos (P>0,05). Estos resultados son similares a los reportados por Alderton et al. (2000). Sin embargo, la diferencia numérica de los grupos suplementados (56 y 50% para el grupo A y B, respectivamente) con respecto al grupo control (25%) es importante. El número de folículos clase 1 y clase 2 durante los primeros cuatro meses del ensayo, no fueron afectados por los tratamientos (P>0,05). Sin embargo, el número de folículos clase 3 fue mayor (P<0,04) en el grupo B que en el grupo control (Figura 7). Estos resultados concuerdan con los datos obtenidos por Grimard et al. (1995), quienes observaron que el aporte nutricional no influyó sobre el número de folículos pequeños y medianos, pero si sobre los folículos de tamaño grande. Estudios previos han sugerido que la adquisición y el mantenimiento subsiguiente de la competencia reproductiva postparto es el resultado de la integración funcional del eje hipotálamo-hipófisis-ovario (Schillo, 1992). Es conocido que durante el periodo postparto se produce una fuerte retroalimentación (feed-back) negativa ejercido por el estradiol a nivel pituitario. Esto trae como consecuencia una disminución en la frecuencia y en la amplitud de los pulsos de LH, que finalmente afectan el crecimiento y el desarrollo folicular. Este efecto negativo del estradiol se ve potenciado por el amamantamiento (Murphy et al., 1990) y por un aporte nutricional inadecuado, el cual es señalizado mediante numerosos signos metabólicos que de una manera integrada influencian los procesos reproductivos. Según Stagg et al. (1995), los procesos involucrados en los eventos tempranos del crecimiento y desarrollo folicular durante el postparto, son independientes de altos o bajos planos nutricionales y del estímulo del amamantamiento, lo cual quizás explica el hecho de no existir diferencias entre tratamientos para las clases foliculares uno y dos. Sin embargo, los eventos finales de maduración folicular requieren de un incremento en la concentración y frecuencia pulsátil de LH (Fortune, 1994), la cual puede estar comprometida por la presencia del becerro, el estímulo del mamado (Williams, 1990), además de la restricción nutricional crónica (Bossis et al., 1999). Investigaciones realizadas por Hess et al. (1998) demostraron que el aporte de aminoácidos (AA) protegidos de la degradación ruminal, incrementa la cantidad de AA disponibles para la gluconeogénesis, además de mantener otras funciones metabólicas como la síntesis proteica. Igualmente, la baja disponibilidad de glucosa, disminuye la liberación hipotalámica de GnRH (Wetteman et al., 2003). Revisiones posteriores realizadas por Hess et al. (2005), sugieren que las regiones del cerebro responsables de la liberación de LH, detectan el incremento en el aporte de los AA metabolizables, o tal vez los imbalances de AA específicos. Estos mismos autores establecen que los múltiples signos metabólicos que reflejan el estatus nutricional son integrados en las regiones del cerebro que regulan la reproducción y que posiblemente ejercen sus efectos al influenciar las hormonas involucradas en la modulación del eje hopotálamo-hipófisis-ovario. En este sentido, Kane et al. (2004) reportaron que la suplementación con PNDR puede alterar la función pituitaria y ovárica, influenciando el comportamiento reproductivo de novillas de carne. Al evaluar el número de animales que presentaron un folículo dominante, se observó que más animales del grupo B (P<0,04) presentaron folículos dominantes, comparados con los animales del grupo control (Figura 8), específicamente durante el primer mes de temporada de servicios. Esta observación coincide con los datos aportados por Grimard et al. (1995), quienes observaron que el número de animales con folículo dominante se incrementa con el tiempo, sobre todo en el grupo de animales que recibían suplementación.
Los resultados obtenidos en el presente ensayo nos permiten inferir que para las vacas de primera lactancia que están restringidas a una temporada de servicios, un manejo apropiado que incluya suplementación estratégica previo al período de servicios, puede mejorar el peso y la condición corporal al inicio de esa temporada, y conjuntamente con la implementación de un sistema de amamantamiento restringido, puede mejorar el comportamiento reproductivo de los animales en esa temporada de servicios. CONCLUSIONES La suplementación con proteína sobrepasante disminuyó el número de días para el primer celo e incrementó la proporción de vacas en celo los primeros sesenta días de temporada de servicios, mientras esos mismos grupos mantenían una ligera ganancia de peso y condición corporal. Este incremento en peso y condición corporal se observa principalmente cuando el forraje es limitado. Una vez que la oferta forrajera aumenta, el efecto no es significativo. Por otra parte, el aumento de la tasa de ganancia de peso y de la condición corporal al inicio de la temporada de servicios está relacionado con el mejoramiento del comportamiento reproductivo, lo cual se traduce en un aumento potencial de vacas preñadas al inicio de la temporada de servicios, un incremento de la vida productiva de la vaca y en una exitosa producción vaca-becerro-año. BIBLIOGRAFÍA Alderton B.W., D.L. Hixon, B.W. Hess, L.F. Woodard, D.M. Hallford y G.E. Moss. 2000. Effects of supplemental protein type on productivity of primiparous cows. J. Anim. Sci., 78: 3027–3035. Anderson L., J. Paterson, R. Ansotegui, M. Cecava y W. Schmutz. 2001. The effects of degradable and undegradable intake protein on the performance of lactating first-calf heifers. J. Anim. Sci., 79: 2224-2232. Bishop D.K., R.P. Wetteman y L.J. Spicer. 1994. Body energy reserves influence the onset of luteal activity after early weaning of beef cows. J. Anim. Sci., 72: 2703. Bossis I., R. Wettemann, S. Welty, J. Vizcarra, L. Spicer y M. Diskin. 1999. Nutritionally induced anovulation in beef heifers: Ovarian and endocrine function preceding cessation of ovulation. J. Anim. Sci., 77: 1536–1546. Ciccioli N., R.P. Wetteman, L.J. Spicer, C.A. Lents, F.J. White y D.H. Keisler. 2003. Influence of body condition at calving and postpartum nutrition on endocrine function and reproductive performance of primiparous beef cows. J. Anim. Sci., 81: 3107-3120. Dhuyvetter D.V., M.K. Petersen, R.P. Ansotegui, R.A. Bellows, B. Nisley, R. Brownson y M.W. Tess. 1993. Reproductive efficiency of range beef cows fed different quantities of ruminally undegradable protein before breeding. J. Anim. Sci., 71: 2586. Díaz T., E.J.P. Schmitt, R.L. De La Sota, M.J. Thatcher y W.W. Thatcher. 1998. Human chorionic gonadotropin-induced alterations in ovarian follicular dynamics during the estrous cycle of heifers. J. Anim. Sci., 76: 1929. Fortune J.E. 1994. Ovarian follicular growth and development in mammals. Biol. Reprod., 50: 225. Grimard B., P. Humblot, A. Ponter, J. Mialot, D. Sauvant y M. Thibier. 1995. Influence of postpartum energy restriction on energy status plasma LH and oestradiol secretion and follicular development in suckled beef cows. J. Reprod. Fertil., 104: 173. Hess B., E.J. Scholljergerdes, S.A. Coleman y J.E. Williams. 1998. Suplemental protein plus ruminally protected methionine and lysine for primiparous beef cattle consuming annual rye hay. J. Anim. Sci., 76:1767-1777. Hess B., S.L. Lake, E.J. Scholljergerdes, T.R. Weston, V. Nayigihugu, J.D. Molle y G.E. Moss. 2005. Nutritional controls of beef cow reproduction. J. Anim. Sci., 83 (Suppl. 1): E90-E106. Kane K., K.W. Creighton, M.K. Petersen, D.M. Hallford, M.D. Remmenga y D.E. Hawkins. 2002. Effects of varying levels of undegradable intake protein on endocrine and metabolic function of young postpartum beef cows. Theriogenology, 57: 2179-2191. Kane K., D.E. Hawkins, G.D. Pulsipher, D.J. Denniston, C.R. Krehbiel, M.G. Thomas, M.K. Petersen, D.M. Hallford, M.D. Remmenga, A.J. Roberts y D.H. Keisler. 2004. Effect of increasing levels of undegradable intake protein on metabolic and endocrine factors in estrous cycling beef heifers. J. Anim. Sci., 82: 283-291. Murphy M.G., M.P. Boland y J.F. Roche. 1990. Pattern of follicular growth and resumption of ovarian activity in postpartum beef suckler cows. J. Reprod. Fertil., 90: 523. Noguchi T. 2000. Protein nutrition and insulin-like growth factor system. Brit. J. Nutr., 84: S241–S244. Nolan C.J., R.C. Bull, R.G. Sasser, C.A. Ruder, P.M. Panlasigui, H.M. Schoenemann y J.J. Reeves. 1988. Postpartum reproduction in protein restricted beef cows: Effect on the hypothalamic- pituitary-ovarian axis. J. Anim. Sci., 66: 3208–3217. NRC (National Research Council). 1996. Nutrient Requirements of Beef Cattle. 7ma ed. National Academy Press, Washington, D.C. Olivares R. 1981. Efecto de la suplementación sobre la eficiencia reproductiva de vacas Brahman de primera lactancia. Tesis de Maestría, Postgrado en Reproducción Animal y Tecnología de la Inseminación Artificial. Facultad de Ciencias Veterinarias, Universidad Central de Venezuela, Maracay, Venezuela. 86p. Plasse D. 1988. Factores que influyen la eficiencia reproductiva de bovinos de carne en América Latina tropical y estrategias para mejorarla. IV Cursillo sobre Bovinos de Carne. Universidad Central de Venezuela, Facultad de Ciencias Veterinarias. Maracay, Venezuela. pp. 1-51. Richards M.W., R.P. Wettemann y H.M. Schoenemann. 1989. Nutritional anestrus in beef cows: Body weight change, body condition, luteinizing hormone in serum and ovarian activity. J. Anim. Sci., 67: 1520–1526. Schillo K.K. 1992. Effects of dietary energy on control of luteinizing hormone secretion in cattle and sheep. J. Anim. Sci., 70: 1271– 1282. Stagg K., M.G. Diskin, J.M. Sreenan y J.F. Roche. 1995. Follicular development in long- term anoestrus suckler beef cows fed two levels of energy postpartum. Anim. Reprod. Sci., 38: 49. Strauch T., E.J. Scholljegerdes, D.J. Patterson, M.F. Smith, M.C. Lucy, W.R. Lamberson y J.E. Williams. 2001. Influence of undegraded intake protein on reproductive performance of primiparous beef heifers maintained on stockpiled fescue pasture. J. Anim. Sci., 79:574-581. Triplett B.L., D.A. Neuendorff y R.D. Randel. 1995. Influence of undegraded intake protein supplementation on milk production, weight gain, and reproductive performance in postpartum Brahman cows. J. Anim. Sci., 73:3223–3229. Wagner J. ., K.S. Lusby, J.W. Oltjen, J. Rakestraw, R.P. Wetteman y L.E. Walters. 1988. Carcass composition in mature hereford cows: estimation and effect on daily metabolizable energy requirements. J. Anim. Sci., 66:603. Wetteman R., C.A. Lents, N.H. Ciccioli, F.J. White y I. Rubio. 2003. Nutritional- and suckling-mediated anovulation in beef cows. J. Anim. Sci., 81(Suppl. 2):E48-E59. Wiley J.S., M.K. Petersen, R.P. Ansotegui y R.A. Bellows. 1991. Production from first-calf beef heifers fed a maintenance or low level of prepartum nutrition and ruminally undegradable or degradable protein postpartum. J. Anim. Sci., 69:4279–4293. Williams G.L. 1990. Suckling as a regulator of postpartum rebreeding in cattle: A review. J. Anim. Sci., 68: 831. Wiltbank M.C., A. Gumen y R. Sartori. 2002. Physiological classification of anovulatory conditions in cattle. Theriogenology, 57:21–52. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||


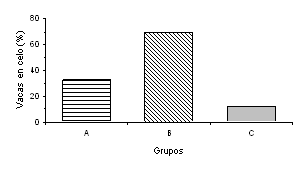
|
|