|
Caña de Azúcar, Vol. 15 (1): 27-37. 1997. CARACTERIZACIÓN
FISIOPATOLOGÍA DE AISLAMIENTOS DEL VIRUS DEL MOSAICO Dolores Piñón*, Ricardo Acevedo*, Omelio Carvajal* y Ondina León** *Instituto
Nacional de Investigaciones de la Caña de Azúcar, |
||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RESUMEN Se realizaron estudios fisiopatológicos con el objetivo de analizar los efectos de distintos aislamientos del SCMV sobre caña de azúcar. Para esto se determinó el porcentaje de infección, cuantificación viral, altura de las plantas, electroforesis de isoperoxidasas, actividad nitratoreductasa y análisis ultraestructurales de tejidos foliares de plantas inoculadas. Se demostró que las variables fisiopatológicas analizadas permitieron diferenciar los aislados de este virus presentes en Cuba. Palabras clave: Virus del mosaico de la caña de azúcar (VMCA), caña de azúcar, análisis de tejido, nitratoreductasa, Elisa, electroforesis, Cuba. PHYSIOPATHOLOGIC
CARACTERIZATION OF SUGARCANE ABSTRACT Physiopathologics studies were carried out to analyze the effects of SCMV isolated in sugarcane plants. Infection percent, viral cuantification, plant height, isoperoxidase electrophoresis, nitratereductase activity and ultrastructural analysis of foliar tissues in inoculated plants were determined. Virus isolates present in Cuba were differentiated by the physiopathologics variables analyzed. Key words: Sugarcane mosaic virus (SCMY), sugarcane, tissue analysis, nitratereductase, Elisa, electrophoresis. INTRODUCCIÓN La enfermedad de la caña de azúcar ocasionada por el virus del mosaico (VMCA) es una de las más ampliamente distribuidas en el mundo. En Cuba se encuentra controlada por el uso de variedades resistentes, pero se continúan los estudios sobre la enfermedad, ya que las condiciones climáticas son favorables para el desarrollo de este virus (10). Han sido reportadas diferentes razas del microorganismo, las que difieren en su habilidad para causar la infección. Los resultados obtenidos más recientemente han demostrado que las razas A, B y D son las más distribuidas mundialmente y que las H e I predominan en Louisiana (6). En Cuba se han reportado las razas A, B y D con predominio de la segunda (5, 10). Estudios más recientes lograron la caracterización de los aislamientos cubanos del VMCA (7), tomando como base no sólo su comportamiento ante los hospedantes diferenciales de caña de azúcar existentes en Cuba, sino también analizando el efecto provocado por éstos en líneas híbridas de sorgo, así como diferentes técnicas serológicas y de microscopía electrónica, determinándose que existe una variabilidad patogénica en el inóculo procedente de la variedad B 34104, pudiendo distinguirse tres aislamientos, cuyas características serológicas, morfológicas y físicas fueron similares. Por todo lo anterior, el objetivo de este trabajo fue la realización de estudios fisiopatológicos que permitieran diferenciar los efectos de estos aislamientos sobre la caña de azúcar. MATERIALES Y MÉTODOS Los aislamientos 2, 3 y 4 del VMCA fueron obtenidos acorde a lo planteado por Carvajal (2), para lo cual se inocularon 80 plantas del hospedente diferencial "CP31-294" con extractos infectivos provenientes de la variedad B 34104. Fueron mantenidas 20 plantas no inoculadas como controles sanos. La infección artificial de las plantas fue por inoculación mecánica empleando el método de tapón (5), cuando las plantas tenían de 2 a 3 hojas. Las plantas fueron cultivadas en casa de cristal, en bolsas de polietileno (46,5 x 37,0 cm3) que contenían suelo Ferralítico Rojo típico, no saturado, previamente esterilizado. El riego fue diario hasta capacidad de campo y la temperatura promedio fue de 36 ± 2° C. A los 28 días posteriores a la inoculación viral, se agruparon las plantas infectadas según los síntomas característicos y descritos para cada uno de los aislamientos (2) y se tomaron muestras aleatorias de las hojas + 1 según Kuijper (12) infectadas con cada aislamiento. La caracterización fisiopatológica se realizó mediante la evaluación de las siguientes variables: - Porcentaje de infección: Sobre la base de la sintomatología típica de la enfermedad. - Cuantificación viral: Se comprobó la ausencia de virus en los controles y se determinó la concentración viral en las plantas infectadas mediante el Test Ultramicroanalítico ELISA (8). - Altura de las plantas: Se midió la longitud en cm de las plantas a partir de la base de la primera hoja emergida hasta el ápice de la hoja más larga. Electroforesis de isoperoxidasas. Se realizó en gel de poliacrilamida al 8,5% con un sistema discontinuo, utilizando como tampón de corrida Tris-glicina pH 8,9 (11). - Actividad nitratoreductasa (NR). La extracción se realizó en tampón fosfato pH 7 con nitrito de potasio 0,1 M. El contenido de nitrito se determinó con el reactivo de Griess (3) tomando como unidad de actividad los micromoles de nitrito formado por gramo de peso fresco por hora. - Análisis ultraestructurales de las hojas. Se fijaron las muestras y se incluyeron en resina Spurr, se cortaron, contrastaron y fueron observadas en el microscopio electrónico (1). Se utilizó un diseño completamente aleatorizado y la evaluación estadística se realizó sobre la base de datos individuales con 6 repeticiones mediante un Análisis de Varianza Simple y la prueba de Newman-Keuls al 5% para la comparación de las medias. RESULTADOS Y DISCUSIÓN El cuadro 1 muestra los resultados obtenidos para las diferentes variables cuantitativas analizadas en las variantes estudiadas. El porcentaje de infección en el aislamiento 3 fue mayor, provocando los síntomas foliares más drásticos en estas plantas, con la casi total destrucción de tejido. En menor proporción apareció el aislamiento 2, cuyos síntomas foliares externos son más leves. Esto coincide con lo repor1ado por otros investigadores (7), corroborando la heterogeneidad del inóculo proveniente de la variedad B 34104. El aislamiento 3 provocó también las mayores reducciones en la altura de las plantas, seguido del aislamiento 4.
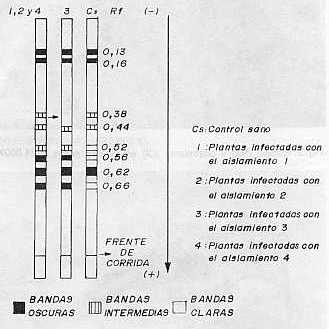
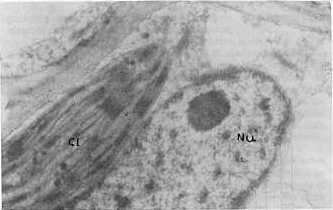
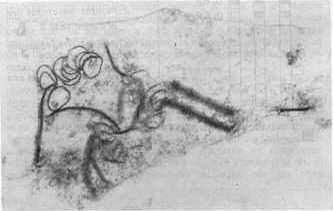
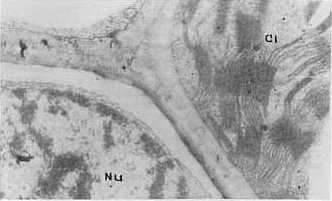
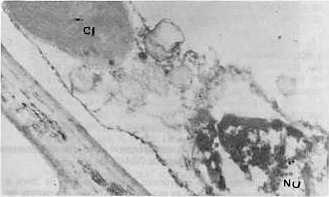
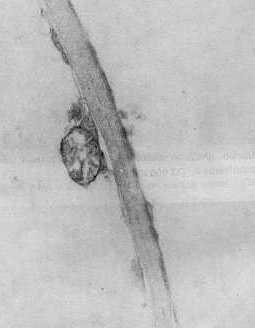
Al analizar la concentración viral, el valor más alto se obtuvo con el aislamiento 2. En orden descendente le siguieron el 4 y el 3, este último con baja concentración. Contrasta el hecho de que siendo el aislamiento 3 el que aparece en mayor porcentaje de infección y el que produce a su vez los daños más notables, sea el de concentración viral más baja a los 28 días de post-inoculación. Sin embargo, es precisamente el aislamiento 3 el que provoca las mayores destrucciones de los tejidos que se observan severamente afectados, por lo cual se hace difícil la sobrevivencia de los virus dada la naturaleza de parásitos obligados y de depender, por tanto, de la maquinaria metabólica de las células para su multiplicación. Estudios de la dinámica de la concentración viral en el aislamiento 2 reflejaron un máximo a los 28 días posteriores a la inoculación (9). Sin embargo los resultados obtenidos en este trabajo hacen pensar en la posibilidad de que el período de máxima concentración viral en el aislamiento 3, se produzca a otros intervalos de tiempo, posiblemente antes de los 28 días de post-inoculación, o que a pesar de no producirse grandes cantidades de virus, éstos sean capaces de ocasionar más afectaciones que concentraciones mayores de otros aislamientos, especialmente del 2. Se detectaron diferencias notables en la actividad NR entre las plantas infectadas con los tres aislamientos, ya que muestras inoculadas con el 2 y el 4 se mantienen con valores superiores al control sano, las enfermas con el aislamiento 3 reflejan muy poca actividad enzimática. Las electroforesis de isoperoxidasas arrojaron que se producía un oscurecimiento intenso en las bandas enzimáticas, para los aislamientos 2 y 4, lo que indica una mayor actividad con respecto a los controles sanos; pero en el caso de las plantas infectadas con el aislamiento 3, se observó la desaparición de una banda con un Rf igual a 0,38 (Fig. 1). En otros sistemas hospedente-parásito se han detectado diferencias en la intensidad de la tinción en las isoenzimas de plantas infectadas con diferentes razas del patógeno (4). Al analizar la ultraestructura se observaron partículas virales típicas de este virus en plantas inoculadas con los tres aislamientos, pero no fueron observadas inclusiones virales en las plantas infectadas con los aislamientos 3 y 4 (Fig. 2 y 3). En el caso del aislamiento 3 se detectaron efectos más drásticos y frecuentes sobre los núcleos y cloroplastos en comparación con los aislamientos 2 y 4 (Fig. 4 y 5). También se observaron mitocondrios con destrucción de su estructura interior (Fig. 6), lo cual no apareció en las plantas afectadas por los otros aislamientos.
La vinculación entre el proceso respiratorio que se lleva a cabo en estos organelos con la actividad PO, nos hace pensar en una posible relación entre las afectaciones mitocondriales observadas y las alteraciones encontradas en la isoperoxidasa de Rf 0,38 en las plantas enfermas con el aislamiento 3 de este virus. CONCLUSIONES Las variables fisiopatológicas: porcentaje de infección, altura, concentración viral y actividad Nitrato Reductasa, permitieron diferenciar sobre la variedad CP 31-294 las plantas infectadas con los aislamientos 2, 3 y 4 del virus del mosaico, mientras que los patrones de isoperoxidasa tuvieron un comportamiento diferencial sólo en el caso del aislamiento 3. El análisis de la ultraestructura celular permitió observar en las plantas infectadas partículas virales típicas del virus del mosaico, así como alteraciones en los cloroplastos, mitocondrios y núcleos que fueron más marcados en el caso del aislamiento 3. También fueron detectadas inclusiones citoplasmáticas en las plantas enfermas con el aislamiento 2. BIBLIOGRAFÍA 1. ACEVEDO, R.; FAJARDO, M. 1988. Optimización de la técnica de inclusión y corte por microscopía electrónica en tejidos foliares de caña de azúcar. Revista INICA, 3: 72-81. 2. CARVAJAL, O.; PERALTA, E.; LEON BLANCO, R.; GONZALEZ, L. ; Regalado, R. 1997. Caracterización y manejo para el control de diferentes aislamientos del VMCA dentro del Programa Nacional de obtención de variedades. Memorias Medio Siglo de Investigaciones Cañeras en Cuba, Jovellanos, 23- 27 Junio, 1997, p 62. 3. ERMAKOV, A.N. 1972. Determinación de la actividad Nitrato Reductasa. En Métodos de Investigaciones Bioquímicas de las Plantas. K. Kolos, Leningrado, 61-62. 4. JENNINGS, P.; BRANNAMAN, B.; ZSHILE, F. 1969. Peroxidase and polyphenoxidase activity associated with Helminthosporium leaf spot of maize. Phytopathol., 59: 963-967. 5. KOLOBAEV, V.; OTERO, I.; MORIN, R.; Carvajal, O. 1974. Propiedades del virus del mosaico de la caña de azúcar. Serie Caña de Azúcar, 46: 1-11. 6. KOIKE, H.; GILLASPIE, A. B. 1989. Mosaic. En Disease of Sugar Cane. Edited by C. Ricaud, B. Egan, a: Gillaspie an C. Hughes, Elsevier Science Publishing. Co., New York, 301-322. 7. LEON, O.; PERALTA, E. L.,; NELSSON, B.; Carvajal, O. 1990. Caracterización de aislamientos y razas del virus del mosaico de la caña de azúcar. II Seminario Científico Internacional de Sanidad Vegetal, 10-14 abril, La Habana, p. 137. 8. PERALTA, E. L.; ALFONSO, R. 1990. Aplicación del sistema ultramicroanalítico (SUMA) al diagnóstico del virus del mosaico mediante el método ELISA-DAS: Evaluación de procedimientos cortos. Memorias I Congreso de la Asociación de Técnicos Azucareros de América Latina y el Caribe, Colombia, 10-14 set., 1990, 143-148. 9. PIÑON, D. 1992. Fisiopatología de la interacción caña de azúcar- virus del mosaico. Tesis presentada en opción al grado científico de Doctor en Ciencias Agrícolas. Instituto Nacional de Investigaciones de la Caña de Azúcar, La Habana, 1992, 110 p. 10. RODRÍGUEZ, E. L. 1979. Fundamentos para los métodos de lucha contra el mosaico de la caña de azúcar en Cuba. Tesis para optar por el Grado Científico de Candidato a Doctor en Ciencias Biológicas. Instituto Pansoviético de Protección de Plantas. Leningrado, 1979, 143 p. 11. RUIZ, A.; MARIBONA, R. 1983. Peroxidase isoenzyme analysis: A massive method for sugarcane varieties identification. Proceeding XVIII Cong. ISSCT, 1: 437-442. 12. VAN DELLEWIJN, C. 1951. Botánica de la caña de azúcar. Pueblo y Educación, La Habana, 1975, 460 p. |
||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||